|
|
|
|
|
2.8 Computerberechnungen von lösungsmittelzugänglichen Oberflächen in Proteinen mit bekannter Struktur |
|
|
2.6 Selektive chemische Modifizierung von Modellproteinen mit Melarsenoxid und massenspektrometrische Charakterisierung der Reaktionsprodukte |
2.7 Charakterisierung der in vitro Rückfaltungsreaktion von rhM-CSF
Das homodimere rhM-CSF (Aminosäuren 4 bis 221) enthält insgesamt 18 Cysteinreste. Die intramolekularen Disulfidbrücken Cys7-Cys90, Cys48-Cys139 und Cys102-Cys146 sitzen außerhalb der -Helices und -Faltblattstrukturen und stabilisieren das Vier-Helix-Bündel. Die intermolekularen Disulfidbrücken Cys31-Cys31, und zusätzlich in der hier zur Verfügung stehenden Form des rhM-CSF, Cys157-Cys157 und Cys159-Cys159 verbinden die beiden Monomereinheiten kovalent zu einem Homodimer mit einem Molekulargewicht von 49030 D. Abb. 78 zeigt die Aminosäuresequenz und die schematisch Disulfidstrukturen von rhM-CSF.

Abb. 78: Aminosäuresequenz für rhM-CSF. Die intramolekularen Disulfidbrücken zwischen Cys7-Cys90, Cys48-Cys139 und Cys102-Cys146 sind durch Striche gekennzeichnet. Die intermolekularen Disulfidbrücken sind an den beteiligten Cysteinen Cys31, Cys157 und Cys159 angedeutet.
Nach Überprüfung der Biscysteinylselektivität von MEL gegenüber vollständig reduziertem bzw. nicht reduziertem rhM-CSF sollte vollständig reduziertes und denaturiertes rhM-CSF zurückgefaltet und die mit IAA und MEL modifizierten Faltungsintermediate massenspektrometrisch und durch SDS-PAGE charakterisiert werden.
2.7.1 Reduktion von rhM-CSF und quantitative Umsetzung mit Melarsen Oxid
Das zur Verfügung stehende rhM-CSF der Herstellerfirma Chiron lag in lyophilisierter Form vor. Nach Solubilisierung wurde das Protein durch Ultrafiltration (siehe 3.6.2) gereinigt und in einen NH4HCO3-Puffer (pH 8.2) überführt. Die Reduktion erfolgte in diesem Puffer mit TCEP (siehe 3.4.1) im molaren Verhältnis 1:180 (Cystein / TCEP 1:10) für 5 h bei 37 C. Ein Aliquot des reduzierten rhM-CSF wurde mit Iodacetamid (siehe 3.4.2) und mittels MALDI-MS-Analyse die vollständige Reduktion und damit neunfache Carboxamidomethylierung des Monomers nachgewiesen. In Abb. 79 A ist zum Vergleich das MALDI-Massenspektrum des rhM-CSF-Homodimers gezeigt. Man erkennt Signale für das einfach bis fünffach protonierte Dimer (D) (m/z 49031, 24515, 16344, 12259 und 9808). Dagegen zeigt das MALDI-Massenspektrum des neunfach carboxamidomethylierten rhM-CSF Monomers ([M(IAA)9]) nur Signale für das einfach bis dreifach protonierte Molekülion (m/z 25038, 12521 und 8349).

Abb. 79: MALDI-Massenspektren (A) des rhM-CSF Homodimers (D) und (B) des vollständig reduzierten und neunfach carboxamidomethylierten rhM-CSF Monomers ([M(IAA)9]).
Reduziertes rhM-CSF wurde mit Melarsen Oxid (MEL) im molaren Verhältnis 3.2:40 (bezogen auf die neun Disulfidbrücken des rhM-CSF-Dimers resultieren molare Verhältnisse von 30:40) in NH4HCO3-Puffer bei pH 8.2 umgesetzt (siehe 3.4.3). Die Reaktion wurde in Gegenwart von 4.5 M Harnstoff durchgeführt, da vollständig MEL-modifiziertes rhM-CSF nach den bisherigen Untersuchungen ohne denaturierende Reagenzien aus der Reaktionslösung ausfällt. Die Reaktion wurde UV-spektroskopisch verfolgt und die Reaktionslösung ohne weitere Reinigungsschritte mittels MALDI-MS untersucht. Die Umsetzung wurde in NH4HCO3-Puffer durchgeführt, obwohl Stabilitätsuntersuchungen an Vas-MEL (siehe 2.4.2) zeigten, daß die Arsonigsäuredithioester in nukleophilen Puffersystemen weniger stabil sind, da so ein direkter Nachweis des MEL-modifizierten rhM-CSF aus der Reaktionslösung durch MALDI-MS (crhm-csf 0.06 µg bzw. 1.6 pmol) möglich war. Ferner zeigten die MALDI-MS-Untersuchungen, daß die Nachweisempfindlichkeit von reduziertem rhM-CSF in 50 mM NaH2PO4 pH 8.5 als Probenpuffer um mindestens den Faktor 10 kleiner ist, als in 10 mM NH4HCO3 pH 8.2. Der zeitliche Verlauf der Absoptionsdifferenz ![]() OD299-267(t) (Abb. 80 B) zeigt, daß die Reaktion innerhalb des Zeitfensters (ca. 20 sec.) zum Öffnen des UV-Photometers beendet ist. Abb. 80 A zeigt das MALDI-Massenspektrum nach 20 min Reaktionszeit. Das aus den einfach bis dreifach protonierten Molekülionen (m/z 8548, 12802, 25631) berechnete durchschnittliche Molekulargewicht beträgt 25625 D. Dies entspricht dem theoretisch berechneten Molekulargewicht von 25620 D für dasvierfach MEL-modifizierte rhM-CSF Monomer und bestätigt die chemische Selektivität von MEL für Biscysteinylgruppen (acht Cysteine). Jedoch lassen sich aus den vorhandenen Daten weder Aussagen darüber treffen, welche der neun Cysteine des rhM-CSF Monomers modifiziert vorliegen, noch welche Cysteinpaare durch MEL verbrückt sind und ob das Reaktionsprodukt heterogen oder homogen vorliegt. Die inAbb. 80 schematisch dargestellten modifizierten Cysteine und die verbrückten Cysteinpaare sind willkürlich gewählt.
OD299-267(t) (Abb. 80 B) zeigt, daß die Reaktion innerhalb des Zeitfensters (ca. 20 sec.) zum Öffnen des UV-Photometers beendet ist. Abb. 80 A zeigt das MALDI-Massenspektrum nach 20 min Reaktionszeit. Das aus den einfach bis dreifach protonierten Molekülionen (m/z 8548, 12802, 25631) berechnete durchschnittliche Molekulargewicht beträgt 25625 D. Dies entspricht dem theoretisch berechneten Molekulargewicht von 25620 D für dasvierfach MEL-modifizierte rhM-CSF Monomer und bestätigt die chemische Selektivität von MEL für Biscysteinylgruppen (acht Cysteine). Jedoch lassen sich aus den vorhandenen Daten weder Aussagen darüber treffen, welche der neun Cysteine des rhM-CSF Monomers modifiziert vorliegen, noch welche Cysteinpaare durch MEL verbrückt sind und ob das Reaktionsprodukt heterogen oder homogen vorliegt. Die inAbb. 80 schematisch dargestellten modifizierten Cysteine und die verbrückten Cysteinpaare sind willkürlich gewählt.
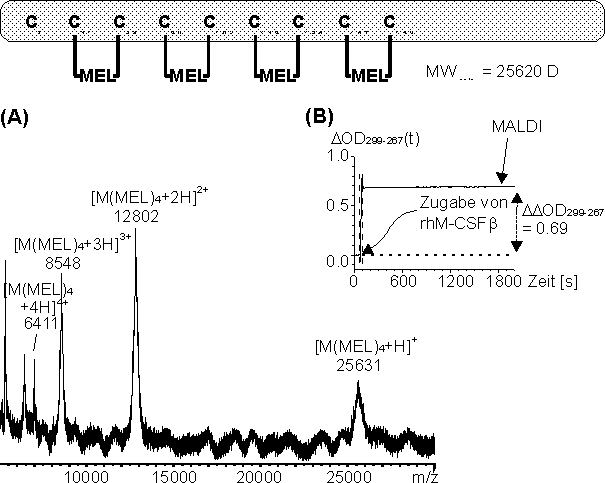
Abb. 80: Umsetzung von vollständig reduziertem rhM-CSF mit MEL im molaren Verhältnis 3.2:40 im NH4HCO3-Puffer bei pH 8.2. (B) MALDI-Massenspektrum der Reaktionslösung nach 30 min Reaktionszeit. (A) Zeitlicher Verlauf der Absorptionsdifferenz ![]() OD299-267(t). Die senkrechten gestrichelten Linien deuten das Öffnen des UV-Photometers an (ca. 20 sec). Des weiteren ist das vierfach MEL-modifizierte rhM-CSF schematisch dargestellt. Die modifizierten Cysteine und die verbrückten Cysteinpaare sind willkürlich gewählt.
OD299-267(t). Die senkrechten gestrichelten Linien deuten das Öffnen des UV-Photometers an (ca. 20 sec). Des weiteren ist das vierfach MEL-modifizierte rhM-CSF schematisch dargestellt. Die modifizierten Cysteine und die verbrückten Cysteinpaare sind willkürlich gewählt.
Die Stabilität der MEL-Modifizierung (siehe 2.4.2) ist eine wesentliche Voraussetzung zur Identifizierung der MEL-modifizierten Biscysteinylfunktionen. Löslichkeitsuntersuchungen [115] zeigten, daß im sauren pH-Bereich (pH 2) das schwerlösliche rhM-CSF Dimer durch Zusatz von Acetonitril (ACN) gut löslich wird. Sowohl das vierfach MEL-modifizierte als auch das vollständig carboxamidomethylierte rhM-CSF Monomer wurden durch Ultrafiltration (siehe 3.6.2) in das Lösungsmittelgemisch, das 40% (v/v) ACN enthielt, umgepuffert und die Retentate MALDI-massenspektrometrisch charakterisiert. Die MALDI-Massenspektren nach Ultrafiltration entsprechen denen in basischen, wäßrigen Puffersystemen (Abb. 79 B undAbb. 80 A).
2.7.2 Reduktion von rhM-CSF mit TCEP in Anwesenheit von Melarsen Oxid und massenspektrometrische Charakterisierung des Reaktionsprodukts
Um die Tauglichkeit der MEL-Abfangreaktion von Faltungsintermediaten für die Proteinfaltung des vollständig reduzierten rhM-CSF zu überprüfen wurde zuerst die reduktive Entfaltung des Dimers untersucht. Die reduktive Entfaltung (local unfolding) des nativ gefaltenen Dimers ist energetisch günstiger, als die komplette Zerstörung der Tertiärstruktur (global unfolding) und kann damit Aussagen über den Mechanismus der Proteinfaltung geben. Zu einer Lösung von MEL in 3 M Harnstoff (NH4HCO3-Puffer, pH 8.2) wurde rhM-CSF Dimer im molaren Verhältnis rhM-CSF / MEL 3.2:40 zugegeben und die Reaktion UV-spektroskopisch verfolgt. Der zeitliche Verlauf der Absorptionsdifferenz OD299-267(t) (sieheAbb. 81 A) zeigt keine Änderung nach Zugabe des Dimers, d.h. MEL reagiert nicht mit Biscystinylgruppen im rhM-CSF (vgl. 2.3.2). Erst nach Zugabe von TCEP im molaren Verhältnis rhM-CSF / TCEP 1:180 bei t 1 h 15 min nimmt ![]() OD299-267(t) zu, d.h. MEL reagiert nun mit den reduziert vorliegenden Cysteinen des rhM-CSF Dimers, und erreicht nach 3-4 h einen Endwert. Das MALDI-Massenspektrum (siehe Abb. 81 B) der Reaktionslösung nach 24 h zeigt Signale bei m/z 49824, 24852, 16627 und 12440 für das einfach bis vierfach protonierte modifizierte Molekülion des rhM-CSF Dimers. Das daraus berechnete Molekulargewicht von 49790 66 D entspricht einer durchschnittlichen 3-fachen MEL-Modifizierung des Dimers. Dies ist in guter Übereinstimmung mit der absoluten Änderung der Absorptionsdifferenz
OD299-267(t) zu, d.h. MEL reagiert nun mit den reduziert vorliegenden Cysteinen des rhM-CSF Dimers, und erreicht nach 3-4 h einen Endwert. Das MALDI-Massenspektrum (siehe Abb. 81 B) der Reaktionslösung nach 24 h zeigt Signale bei m/z 49824, 24852, 16627 und 12440 für das einfach bis vierfach protonierte modifizierte Molekülion des rhM-CSF Dimers. Das daraus berechnete Molekulargewicht von 49790 66 D entspricht einer durchschnittlichen 3-fachen MEL-Modifizierung des Dimers. Dies ist in guter Übereinstimmung mit der absoluten Änderung der Absorptionsdifferenz ![]() OD299-267 zwischen der Zugabe des Reduktionsmittels und dem Erreichen des Plateaus (Abb. 81 A: 1 h 15 min t 5 h). Diese ist mit 0.24 deutlich kleiner als für die Umsetzung von vollständig reduzierten rhM-CSF mit MEL (OD299-267 0.69, sieheAbb. 80 A). Der Wert
OD299-267 zwischen der Zugabe des Reduktionsmittels und dem Erreichen des Plateaus (Abb. 81 A: 1 h 15 min t 5 h). Diese ist mit 0.24 deutlich kleiner als für die Umsetzung von vollständig reduzierten rhM-CSF mit MEL (OD299-267 0.69, sieheAbb. 80 A). Der Wert ![]() OD299-267 0.69 entspricht acht MEL-Modifizierungen (bezogen auf das Dimer); somit errechnet man für OD299-267 0.24 eine durchschnittliche 2.7-fache MEL-Modifizierung des rhM-CSF Dimers.
OD299-267 0.69 entspricht acht MEL-Modifizierungen (bezogen auf das Dimer); somit errechnet man für OD299-267 0.24 eine durchschnittliche 2.7-fache MEL-Modifizierung des rhM-CSF Dimers.
Die unvollständige Reduktion kann nicht auf die Reduktionsbedingungen zurückgeführt werden, da die verwendeten molaren Verhältnisse an rhM-CSF zu TCEP, sowie Temperatur und Reaktionszeit nahezu identisch zu früheren, vergleichbaren Reaktionsansätzen ohne MEL-Anwesenheit (siehe 2.7.1) waren, in denen jedoch rhM-CSF quantitativ reduziert wurde. Untersuchungen der reduktiven Entfaltung (reductive unfolding) an Ribonuklease A (RNAse A) [148] zeigen, daß sich während der Reduktion von RNAse A Intermediate anreichern, die drei der vier nativen Disulfidbrücken enthalten. Der geschwindigkeitsbestimmende Schritt ist dann die weitere Reduktion der noch intakten nativen Disulfidstrukturen. In gleicher Weise kann für die durchgeführte reduktive Entfaltung von rhM-CSF, in Gegenwart des 'Trapping'-Reagenz MEL, eine bevorzugte Reduktion bestimmter Disulfidbrücken erfolgen. Oxidative Faltungsstudien am verkürzten rhM-CSF (aa 4-154) [118], sowie am rhM-CSF [37] zeigten, daß sich die Disulfidbrücken Cys102-Cys146 und Cys31-Cys31 erst spät in der in vitro Faltung schließen. Erst nach Ausbildung der Cys102-Cys146 Disulfidbrücke besitzt das rhM-CSF Dimer seine hohe Resistenz gegenüber Proteasen [37], da so die sehr kompakte Struktur des Vier-Helix-Bündels stabilisiert wird. Für die in vivo Faltung ist eine späte Ausbildung von Disulfidbrücken, die dem Protein proteolytische Resistenz verleihen sinnvoll. Denkbar wäre, daß umgekehrt bei der reduktiven Entfaltung die Disulfidbrücken Cys102-Cys146 und Cys31-Cys31 mit als erste reduziert werden.
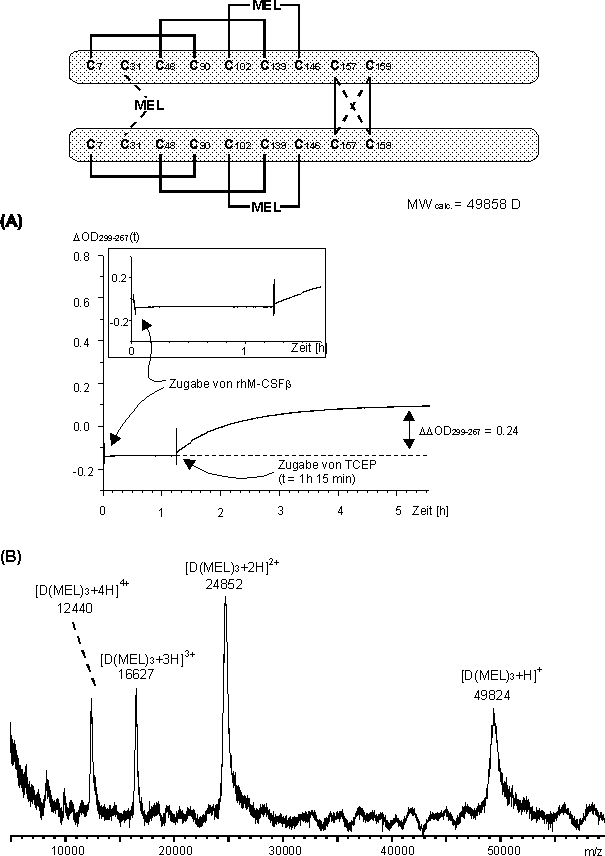
Abb. 81: In situ-Reduktion des rhM-CSF Dimer und Derivatisierung mit MEL im NH4HCO3-Puffer bei pH 8.2 im molaren Verhältnis rhM-CSF /TCEP / MEL 3.2:600:40. (A) Zeitlicher Verlauf der Absorptionsdifferenz ![]() OD299-267(t). Der Ausschnitt (t < 1.5 h) zeigt, daß nach Zugabe des rhM-CSF Dimers keine zeitliche Änderung von
OD299-267(t). Der Ausschnitt (t < 1.5 h) zeigt, daß nach Zugabe des rhM-CSF Dimers keine zeitliche Änderung von ![]() OD299-267(t) eintritt. (B) MALDI-Massenspektrum der Reaktionslösung nach 24 h Reaktionszeit. Schematisch ist die vorgeschlagene Struktur des dreifach MEL-modifizierten rhM-CSF Dimers dargestellt .
OD299-267(t) eintritt. (B) MALDI-Massenspektrum der Reaktionslösung nach 24 h Reaktionszeit. Schematisch ist die vorgeschlagene Struktur des dreifach MEL-modifizierten rhM-CSF Dimers dargestellt .
Die weitere Entfaltung und fortschreitende Reduktion könnte dadurch verhindert werden, daß MEL sofort mit den entstandenen Biscysteinylfunktionen zu den entsprechenden Arsonigsäuredithioestern reagiert und somit die "native" Struktur des rhM-CSF Dimers weitgehend erhalten bleibt. Die schematische Darstellung des dreifach MEL-modifizierten rhM-CSF Dimers (siehe Abb. 81) verdeutlicht die vorgeschlagene symmetrische Einfachmodifizierung jeweils eines Monomers und die verbrückende kovalente Modifizierung zwischen den Monomerhälften.
2.7.3 Charakterisierung des Reaktionsverlaufs der in vitro Rückfaltungsreaktion durch Molekulargewichtsbestimmung der Faltungsintermediate
Auf das Prinzip der oxidativen Rückfaltungsreaktion wurde bereits in Abschnitt 1.1.2 eingegangen. Die Rückfaltung und Modifizierung der Faltungsintermediate von rhM-CSF erfolgte in Gegenwart von GSH; deshalb mußte untersucht werden, inwieweit GSH die Bildung von zyklischen Arsonigsäuredithioestern aus MEL und Proteindithiolen beeinflußt.
Reaktion von MEL mit Oxytocin in Gegenwart von reduziertem Glutathion
Wie in Abschnitt 2.2.2 beschrieben reagiert das Monothiol Glutathion (GSH) mit MEL zu dem entsprechenden offenkettigen Arsonigsäuredithioester. MEL wurde im NaH2PO4-Puffer bei pH 8.5 mit GSH im molaren Verhältnis MEL / GSH 4:12.5 versetzt. Die Reaktion wurde UV-spektroskopisch und MALDI-massenspektrometrisch verfolgt. Die Zunahme ![]() OD299-267(t) (siehe Abb. 82 A) nach Zugabe von GSH zu MEL zeigt die Bildung des offenkettigen Arsonigsäurediesters an. Das MALDI-Massenspektrum (sieheAbb. 82 B) der Reaktionslösung zum Zeitpunkt t = 2400 s zeigt neben MEL (m/z 311) und seinen Mono- und Diester mit der Matrix HCCA (m/z 482 und 653, vgl. Abb. 30) Signale für den offenkettigen Dithioester MEL-(SG)2 (m/z 889, Na-Addukt: 911). Das Signal bei m/z 582 könnte dem Monothioester entsprechen, unter der Voraussetzung, daß der N-Terminus von GSH an der Ausbildung einer zyklischen Struktur durch Kondensation (Verlust von Wasser, 18 amu) beteiligt ist.
OD299-267(t) (siehe Abb. 82 A) nach Zugabe von GSH zu MEL zeigt die Bildung des offenkettigen Arsonigsäurediesters an. Das MALDI-Massenspektrum (sieheAbb. 82 B) der Reaktionslösung zum Zeitpunkt t = 2400 s zeigt neben MEL (m/z 311) und seinen Mono- und Diester mit der Matrix HCCA (m/z 482 und 653, vgl. Abb. 30) Signale für den offenkettigen Dithioester MEL-(SG)2 (m/z 889, Na-Addukt: 911). Das Signal bei m/z 582 könnte dem Monothioester entsprechen, unter der Voraussetzung, daß der N-Terminus von GSH an der Ausbildung einer zyklischen Struktur durch Kondensation (Verlust von Wasser, 18 amu) beteiligt ist.
Zu dieser Lösung wurde zum Zeitpunkt t = 2400 s TCEP-reduziertes Oxytocin (OT) zugegeben, so daß ein molares Verhältnis MEL / GSH / OT 5:12.5:3 resultierte. Die Absorptionsdifferenz ![]() OD299-267(t) zeigt einen deutlichen raschen Anstieg (siehe Abb. 82 A) und erreicht einen konstanten Endwert innerhalb des Zeitfensters zum Öffnen des UV-Photometers. Das MALDI-Massenspektrum (siehe Abb. 82 B) zum Zeitpunkt t = 2500 s zeigt nahezu vollständig MEL-modifiziertes Oxytocin (OT-MEL) bei m/z 1284 (Na-Addukt: 1306), neben geringen Anteilen an nicht modifizierten OT (Na-Addukt 1029). Trotz des zweifach höheren molaren Überschusses an GSH bezogen auf die freie Proteinthiolkonzentration bestätigt die Ausbildung des OT-MEL-Addukts die höhere Stabilität zyklischer Arsonigsäuredithioester gegenüber offenkettigen. Dies ist eine wichtige Voraussetzung zur Charakterisierung von Faltungsintermediaten mit MEL in Gegenwart von GSH.
OD299-267(t) zeigt einen deutlichen raschen Anstieg (siehe Abb. 82 A) und erreicht einen konstanten Endwert innerhalb des Zeitfensters zum Öffnen des UV-Photometers. Das MALDI-Massenspektrum (siehe Abb. 82 B) zum Zeitpunkt t = 2500 s zeigt nahezu vollständig MEL-modifiziertes Oxytocin (OT-MEL) bei m/z 1284 (Na-Addukt: 1306), neben geringen Anteilen an nicht modifizierten OT (Na-Addukt 1029). Trotz des zweifach höheren molaren Überschusses an GSH bezogen auf die freie Proteinthiolkonzentration bestätigt die Ausbildung des OT-MEL-Addukts die höhere Stabilität zyklischer Arsonigsäuredithioester gegenüber offenkettigen. Dies ist eine wichtige Voraussetzung zur Charakterisierung von Faltungsintermediaten mit MEL in Gegenwart von GSH.
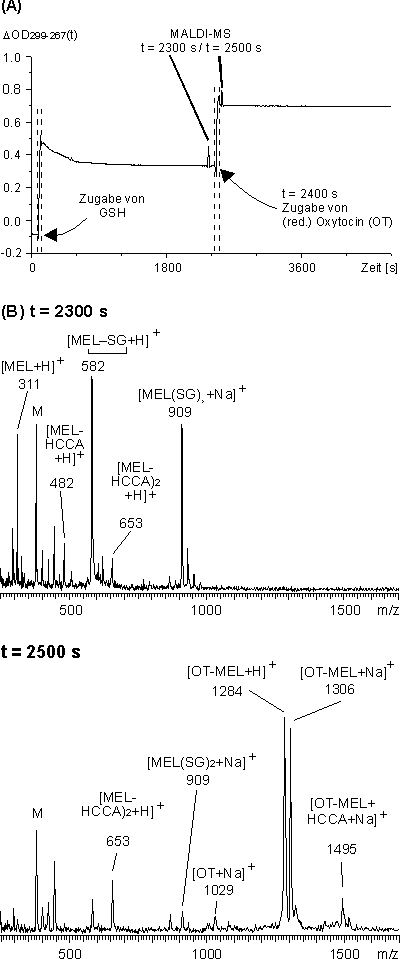
Abb. 82: (A) Zeitlicher Verlauf der Absorptionsdifferenz ![]() OD299-267(t) für die Umsetzung von MEL im NaH2PO4-Puffer (pH 8.5) mit GSH im molaren Verhältnis 4:12.5. Bei t = 2400 s wurde TCEP-reduziertes Oxytocin zugegeben, so daß ein molares Verhältnis MEL / GSH / OT 5:12.5:3 resultierte. (B) MALDI-Massenspektren der Reaktionslösung zum Zeitpunkt t = 2300 s und t = 2500 s. 'M' entsprechen Signale der Matrix HCCA.
OD299-267(t) für die Umsetzung von MEL im NaH2PO4-Puffer (pH 8.5) mit GSH im molaren Verhältnis 4:12.5. Bei t = 2400 s wurde TCEP-reduziertes Oxytocin zugegeben, so daß ein molares Verhältnis MEL / GSH / OT 5:12.5:3 resultierte. (B) MALDI-Massenspektren der Reaktionslösung zum Zeitpunkt t = 2300 s und t = 2500 s. 'M' entsprechen Signale der Matrix HCCA.
Oxidative Rückfaltung von rhM-CSF mit DDT als Reduktionsmittel
Das lyophilisiert vorliegende rhM-CSF Dimer wurde vollständig mit DTT reduziert und durch Harnstoff denaturiert (siehe 3.8) [37]. Die Vollständigkeit der Reduktion wurde durch die Umsetzung mit Iodacetamid und MALDI-MS-Analyse bestätigt (vgl.Abb. 80 B). Die Rückfaltung wurde durch 30-faches Verdünnen des vollständig reduzierten und denaturierten Monomers in einen NH4HCO3-Puffer (pH 8.5) mit hohem Reduktionspotential (0.4 mM GSH, 0.2 mM GSSG) initiiert. Das hohe Reduktionspotential [( 0.34 mM DTT- bzw. Protein-SH plus 0.39 mM GSH ) / 0.38 mM GS- 1.9] sollte die mögliche Oxidation des Monomers vermindern [37]. Die resultierende Proteinkonzentration cm-csf 0.5 µg/µl sollte während der Rückfaltungsreaktion nicht überschritten werden, da sonst ein Großteil des Proteins in Form kovalent aggregierter Monomere vorliegt und sich somit der weiteren Charakterisierung entzieht [112]. Aus dem gleichen Grund wurde die Rückfaltung bei 4 C durchgeführt. Nach 0 h (also direkt nach dem Start der Rückfaltungsreaktion), 0.25 h, 1 h, 3 h, 6 h, 18 h und 42 h wurden je zwei Aliquote entnommen und mit Iodacetamid (IAA) bzw. Melarsen Oxid (MEL) umgesetzt (siehe 3.8). Die Carboxamidomethylierung erfolgte im molaren Verhältnis Thiol / IAA 1:20 bezogen auf die gesamt vorliegende Thiolkonzentration (reduziertes DTT, GSH und Proteinthiole). Die Umsetzung mit MEL wurde im NH4HCO3-Puffer bei pH 8.2 in Gegenwart von 3 M Harnstoff durchgeführt, um Löslichkeitsprobleme der MEL-modifizierten rhM-CSF Rückfaltungsintermediate auszuschließen. Die Bedingungen wurden so gewählt, daß MEL im 10%igen molaren Überschuß zu den freien Thiolen (DTT und M-CSF-Cysteine) vorlag. Wegen der hohen DTT-Konzentration und der damit verbundenen Bildung des MEL-DTT-Addukts wurde auf die UV-spektroskopische Kontrolle des Reaktionsverlaufs verzichtet. Die Reaktion wurde nach einer Reaktionszeit von 10 min als beendet angesehen, aufgrund der an den Modellpeptiden und
-proteinen beobachteten hohen Reaktionsgeschwindigkeit der MEL-Modifizierung bei pH 8 (siehe 2.2.3 und 2.6). Die Reaktionslösungen für beide Modifizierungsansätze wurden unmittelbar nach der Modifizierung eingefroren, um hydrolytische Spaltungen (siehe 2.4.2) der M-CSF-MEL-Addukte durch GSH bei pH 8.2 zu minimieren. Ohne weitere Reinigung erfolgte eine erste Charakterisierung der so erhaltenen Faltungsintermediate durch SDS-PAGE (s.u.) und MALDI-MS. Anschließend wurden Aliquote der mit IAA und MEL modifizierten rhM-CSF Faltungsintermediate durch Ultrafiltration von den überschüssigen Reagenzien abgetrennt und in das Lösungsmittelgemisch 0.1% TFA / 0.1% TFA in ACN 6:4 (pH 2) umgepuffert, wodurch gleichzeitig eine Stabilisierung der MEL-Modifizierung erfolgte (siehe 2.4.2). Die anschließende MALDI-MS-Analyse zeigte vergleichbare Resultate zu dem MALDI-Massenspektren vor der Ultrafiltration.
In Abb. 83 A ist das SDS-Polyacrylamid Gel (SDS-PAGE) unter nicht reduzierenden Bedingungen dercarboxamidomethylierten Faltungsintermediate (vor der Ultrafiltration) für die Zeitpunkte 0 h -42 h dargestellt. Unmittelbar nach dem Start der Rückfaltung (0 h) bis zum Zeitpunkt t = 1 h liegt das rhM-CSF überwiegend als Monomer vor. Dabei läuft die Bande für t = 0 bis 1 h deutlich höher (ca. 32 kD) als die des reduzierten, aber nicht carboxamidomethylierten rhM-CSF Monomers (Spur M) (ca. 28 kD). Im weiteren zeitlichen Verlauf der Rückfaltung nimmt der Anteil an Monomer ab. Die geringen Anteile an monomeren rhM-CSF zeigen nun ein ähnliches Laufverhalten wie das reduzierte rhM-CSF Monomer (Spur M). Nach ca. 6 h liegt praktisch ausschließlich das dimere rhM-CSF (Spur 6 h bis 42 h) vor. Die MALDI-Massenspektren der entsprechenden carboxamidomethylierten Faltungsintermediate nach Ultrafiltration sind inAbb. 83 B gezeigt. Die in eckigen Klammern angegebenen Zahlen entsprechen den durchschnittlich eingeführten Carboxamidomethylierungen bezogen auf das rhM-CSF Dimer. Dargestellt ist nur der Ausschnitt für das einfach protonierte rhM-CSF Dimer [D+H]+ und für das zweifach protonierte Dimer [D+2H] 2+, das mit dem einfach protonierten Monomer [M+H]+ überlagert. Zur Berechnung des durchschnittlichen Molekulargewichts und damit des durchschnittlichen Modifizierungsgrades (siehe Tab. 8) wurden die entsprechenden dreifach protonierten Molekülionen des Monomers und die dreifach und vierfach protonierten Molekülionen des Dimers mit einbezogen. Das MALDI-MS zum Zeitpunkt t = 0 h zeigt ausschließlich das neunfach carboxamidomethylierte rhM-CSF Monomer. Der zeitliche Verlauf zeigt die Zunahme der Signalintensität des Dimers (m/z 50 kD) in Analogie zur SDS-PAGE (Abb. 83 A). Als zusätzliche Information erhält man die Abnahme des durchschnittlichen Modifizierungsgrades (Abb. 83 B). Nach 42 h liegt das rhM-CSF überwiegend als nicht modifiziertes Dimer vor; damit ist die Rückfaltungsreaktion beendet. Die Anzahl der durchschnittlichen Modifizierungsgrade der untersuchten Faltungsintermediate steht in guter Übereinstimmung mit früheren MS-Untersuchungen [37].

Abb. 83: (A) SDS-PAGE der Rückfaltungsreaktion von DTT-reduziertem rhM-CSF. Die Faltungsintermediate nach 0 h bis 42 h wurden durch Carboxamidomethylierung stabilisiert. Aufgetragen sind die Reaktionslösungen vor der Ultrafiltration (entsprechend 1 µg). Mit LMW ist der Molekulargewichtsmarker gekennzeichnet. M entspricht dem reduzierten, aber nicht carboxamidomethylierten rhM-CSF Monomer (2 µg); D dem intakten rhM-CSF Dimer (2 µg). (B) MALDI-Massenspektren der carboxamidomethylierten Faltungsintermediate nach 0 h bis 42 h nach Ultrafiltration. Dargestellt ist der Ausschnitt für das einfach protonierte rhM-CSF Dimer [D+H]+ und für das zweifach protonierte Dimer [D+2H] 2+, das mit dem protonierten Monomer [M+H]+ überlagert. Die Zahlen in den eckigen Klammern geben die Anzahl der eingeführten Carboxamidomethylierungen bezogen auf das rhM-CSF Dimer an. Die entsprechenden m/z-Werte sind in Tab. 8zusammengefaßt.
Die in vitro Rückfaltungsreaktion wurde wie oben beschrieben parallel zur IAA-Alkylierung der Faltungsintermediate, zusätzlich mit MEL als Abfangreagenz durchgeführt und die MEL-modifizierten Faltungsintermediate mit SDS-PAGE und MALDI-MS analysiert. Die inAbb. 84 A dargestellte SDS-PAGE-Untersuchung (unter nicht reduzierenden Bedingungen) der MEL-modifizierten Faltungsintermediate vor der Ultrafiltration unterscheidet sich deutlich zu der oben beschriebenen Faltungsreaktion mit IAA (Abb. 83 A): Der Anteil an rhM-CSF Monomer ist in allen Spuren deutlich höher als bei den mit IAA modifizierten Faltungsintermediaten; umgekehrt ist der Anteil an gebildetem Dimer geringer. Anders als bei der IAA-Modifizierung, liegen in der SDS-PAGE-Analyse der MEL-Derivate wohl keine modifizierten Proteine vor, da durch das 5-minutige Erhitzen (95 C) der Probe (siehe 3.6.4) im Gelaufschlußpuffer (pH > 7) z.T. die erneute Reduktion des gebildeten rhM-CSF Dimers durch noch vorhandenes GSH, sowie eine partielle Hydrolyse der MEL-Modifizierung aufgrund der erhöhten Temperatur und des pH-Werts auftritt. Unter diesen Bedingungen könnte zusätzlich die Spaltung der M-CSF-MEL-Addukte durch GSH erfolgen. Dies wird durch die auf der gleichen Höhe (ca. 29 kD) laufenden Banden des rhM-CSF Monomers zu Beginn der Rückfaltungsreaktion (Spur 0 h, 0.25 h und 1 h in Abb. 84 A, vgl. dazu die gleichen Spuren in Abb. 83) nahegelegt. Dagegen spielen Hydrolyseprozesse bei der MALDI-MS-Analyse (Abb. 84 B) der MEL-modifizierten Faltungsintermediate nach Ultrafiltration und Umpuffern auf pH 2 (s.o.) keine Rolle. Die gefunden durchschnittlichen MEL-Modifizierungen (von Biscysteinylfunktionen) stehen in guter Übereinstimmung zu den Ergebnissen der IAA-Alkylierung (vgl. Abb. 83 B und Tab. 9). Dabei entspricht die achtfache MEL-Modifizierung bezogen auf das rhM-CSF Dimer dem vierfach MEL- und damit vollständig modifizierten rhM-CSF Monomer. Der zeitliche Verlauf zeigt auch hier die Abnahme der durchschnittlichen MEL-Modifizierung bis zum nicht modifizierten, vollständig renaturierten rhM-CSF Dimer nach 42 h.
Zu Beginn der Rückfaltungsreaktion liegt das rhM-CSF vollständig reduziert vor. Die entsprechenden IAA oder MEL modifizierten Faltungsintermediate enthalten daher eine einheitliche Modifizierung; die MALDI-MS-Signale sind dementsprechend schmal. Im Verlauf der Rückfaltung bilden sich zu unterschiedlichen Anteilen Faltungsintermediate mit unterschiedlicher Anzahl an Disulfidbrücken. Die daraus folgende Heterogenität der Modifizierungen führt zu Signalverbreiterungen in den MALDI-Massenspektren. Für das am Ende der Rückfaltung einheitlich erhaltene native rhM-CSF Dimer erhält man wieder deutlich schmalere Halbwertsbreiten der MALDI-MS-Signale. Die beobachteten Peakverbreiterungen geben, insbesondere bei der Modifizierung mit MEL (Abb. 84 B, Spur 1 h bis 6 h), diese Heterogenität wieder. Die unterschiedliche Anzahl eingeführter MEL-Modifizierungen (M = 276) lassen sich selbst am zweifach und dreifach protonierten Molekülions des rhM-CSF Monomers unterscheiden. Dagegen können die geringen Massenunterschiede von m = 58 pro eingeführter Carboxamidomethylierung, aufgrund des hohen Molekulargewichts des rhM-CSF Monomers bzw. Dimers, kaum aufgelöst werden. In Abb. 85 ist dies am Beispiel der Rückfaltungsintermediate nach 3 h dargestellt. Während der Ausschnitt des MALDI-Massenspektrums inAbb. 85 A nur die Aussage einer durchschnittlich 4.4-fachen Carboxamidomethylierung zuläßt, kann inAbb. 85 B genau zwischen den einzelnen ein- bis vierfachen MEL-Modifizierungen unterschieden werden.

Abb. 84: (A) SDS-PAGE der Rückfaltungsreaktion von DTT-reduziertem rhM-CSF. Die Faltungsintermediate nach 0 h bis 42 h wurden durch MEL-Modifizierung stabilisiert. Aufgetragen sind die Reaktionslösungen vor der Ultrafiltration (entsprechend 1 µg). Mit LMW ist der Molekulargewichtsmarker gekennzeichnet. M entspricht dem reduzierten, aber nicht modifizierten rhM-CSF Monomer (2 µg); D dem intakten rhM-CSF Dimer (2 µg). (B) MALDI-Massenspektren der MEL-modifizierten Faltungsintermediate nach 0 h bis 42 h nach Ultrafiltration. Dargestellt ist der Ausschnitt für das einfach protonierte rhM-CSF Dimer [D+H]+ und für das zweifach protonierte Dimer [D+2H] 2+, das mit dem einfach protonierten Monomer [M+H]+ überlagert. Die Zahlen in den eckigen Klammern geben die Anzahl der eingeführten MEL-Modifizierungen bezogen auf das rhM-CSF Dimer an. Die entsprechenden m/z-Werte sind in Tab. 9zusammengefaßt.
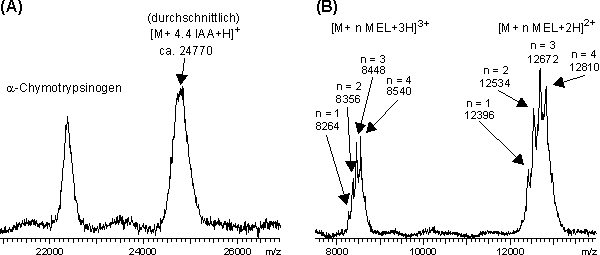
Abb. 85: Dargestellt ist die durch MALDI-MS nachgewiesene Heterogenität der (A) carboxamidomethylierten und(B) MEL-modifizierten Faltungsintermediate nach 3 h. Das Spektrum (A) wurde intern mit -Chymotrypsinogen kalibriert.
Das durchschnittliche Molekulargewicht der carboxamidomethylierten rhM-CSF Faltungsintermediate wurde aus den centroidierten Signalen der MALDI-Massenspektren für die einfach bis vierfach protonierten Molekülionen berechnet (Tab. 8, Spalte: m/z). Dagegen wurde das durchschnittliche Molekulargewicht der MEL-modifizierten rhM-CSF Faltungsintermediate pro Modifizierungsgrad berechnet (Tab. 9, Spalte m/z), da die unterschiedliche Anzahl der eingeführten MEL-Moleküle für die einfach bis vierfach protonierten Molekülionen unterschieden werden konnte (siehe Abb. 85). Aus den so ermittelten durchschnittlichen Molekulargewichten wurde die Anzahl der eingeführten Modifizierungen (Spalte n) bzw. der bereits ausgebildeten Disulfidbrücken (Spalte m) bezogen auf das rhM-CSF Dimer abgeschätzt. In Tab. 8 bzw.Tab. 9 sind die erhaltenen Werte für die carboxamidomethylierten bzw. MEL-modifizierten Faltungsintermediate zusammengefaßt.
Tab. 8: MALDI-MS Molekulargewichtsbestimmungen der carboxamidomethylierten Faltungsintermediate des mit DTT reduzierten rhM-CSF (nach Ultrafiltration). Die m/z-Werte geben das durchschnittlich Molekulargewicht an (siehe Text). Die Werte für n(IAA) bzw. m(SS) geben die Anzahl der eingeführten Carboxamidomethylierungen bzw. der bereits ausgebildeten Disulfidbrücken bezogen auf das Dimer an.
|
[DH2]2+ / [MH]+ |
[DH]+ |
|||||
|
zeitlicher |
m/z(Durchschn.) |
n(IAA) bez. auf Dimer |
m(SS) bez. auf Dimer |
m/z (Durchschn.) |
n(IAA) bez. auf Dimer |
m(SS) bez. auf Dimer |
|
0 h |
25037 |
18 |
0 |
- |
- |
- |
|
0.25 h |
25026 |
17.6 |
0 |
49818 |
13.6 |
2 |
|
1 h |
24914 |
13.8 |
2 |
49868 |
14.4 |
2 |
|
3 h |
24770 |
8.8 |
4-5 |
49527 |
8.6 |
4-5 |
|
6 h |
24651 |
4.6 |
6-7 |
49311 |
4.8 |
6-7 |
|
18 h |
24561 |
1.6 |
8-9 |
49047 |
0.3 |
8-9 |
|
42 h |
24532 |
0.6 |
8-9 |
49026 |
0 |
9 |
Tab. 9: MALDI-MS Molekulargewichtsbestimmungen der MEL-modifizierten Faltungsintermediate pro Modifizierungsgrad (siehe Text) des mit DTT reduzierten rhM-CSF (nach Ultrafiltration). Die Werte für n(MEL) bzw. m(SS) geben die Anzahl der eingeführten MEL-Modifizierungen bzw. der bereits ausgebildeten Disulfidbrücken bezogen auf das Dimer an.
|
[DH2]2+ / [MH]+ |
[DH]+ |
|||||
|
zeitlicher Verlauf |
m/z(Durchschn.) |
n(MEL) bez. auf Dimer |
m(SS) bez. auf Dimer |
m/z (Durchschn.) |
n(MEL) bez. auf Dimer |
m(SS) bez. auf Dimer |
|
0 h |
25651 |
8.2 |
0 |
- |
- |
- |
|
0.25 h |
25668 |
8.2 |
0 |
- |
- |
- |
|
1 h |
25625 |
8 |
0 |
50921 |
6.9 |
2 |
|
25358 |
6.2 |
2-3 |
||||
|
3 h |
25296 |
5.6 |
2-3 |
50736 |
6.2 |
2-3 |
|
25111 |
4.4 |
4-5 |
||||
|
6 h |
25141 |
4.6 |
4-5 |
48147 |
2.7 |
6-7 |
|
24843 |
2.4 |
6-7 |
||||
|
18 h |
24820 |
3.3 |
5-6 |
49689 |
2.4 |
6-7 |
|
42 h |
24603 |
0.6 |
8-9 |
49116 |
0.3 |
9 |
Oxidative Rückfaltung von rhM-CSF mit TCEP als Reduktionsmittel
In einer weiteren Versuchsreihe wurde das Reduktionsmittel DTT durch TCEP ersetzt (siehe 3.8). Die vollständige Reduktion wurde durch IAA-Alkylierung und MALDI-MS-Analyse (vgl. Abb. 80 B) bestätigt. Die Rückfaltung wurde analog dem oben beschriebenen Verfahren durch 30-faches Verdünnen des vollständig reduzierten und denaturierten rhM-CSF Monomers in einen NH4HCO3-Puffer (pH 8.5), der 0.4 mM GSH und 0.4 mM GSSG enthielt, initiiert. Das thiolfreie Reduktionsmittel TCEP ist nicht in den Rückfaltungsgleichgewichten (siehe Abb. 1) involviert. Es wurde angenommen, daß das nach der Reduktion des rhM-CSF Dimers überschüssige TCEP unmittelbar nach dem Start der Rückfaltungsreaktion das zugegebene GSSG zu GSH reduziert. Aus diesem Grund wurde die GSSG Konzentration im Rückfaltungspuffer auf 0.4 mM erhöht. Das unter diesen Bedingungen berechnete Reduktionspotential ( 0.18 mM Proteinthiole + 0.53 mM GSH) / 0.58 GS-) beträgt 1.2. Nach 0 h, 0.25 h, 1 h, 3 h, 6 h, 18 h und 42 h wurden Aliquote der Rückfaltungslösung entnommen und mit IAA und MEL umgesetzt. Die IAA-Alkylierung erfolgte im molaren Verhältnis Thiol (= Protein-SH + GSH) / IAA 1:35. Die Umsetzung mit MEL in Gegenwart von 3 M Harnstoff erfolgte im molaren Verhältnis Biscysteinylfunktionen (= M-CSF-thiole) zu MEL von 3.7:6, der molare Überschuß an MEL betrug also 62 %. Die Bildung der nicht stabilen offenkettigen Arsonigsäuredithioester aus MEL und GSH wurden für die Modifizierungsreaktion nicht berücksichtigt (s.o.). Auch hier wurde die Reaktion nach 10 min (25 C) als beendet angesehen und anschließend, um Hydrolyseeffekte (s.o.) zu vermeiden, sofort tiefgefroren. Die Charakterisierung der so modifizierten Faltungsintermediate erfolgte vorUltrafiltration und Umpuffern auf pH 2 (s.o.) durch SDS-PAGE und MALDI-MS-Analysen. Die MALDI-MS-Analyse nach Ultrafiltration zeigte vergleichbare Spektren.
Die Abb. 86 A dargestellte SDS-PAGE (nicht reduzierend) der carboxamidomethylierten Faltungsintermediate für die Zeitpunkte 0 h bis 42 h ist im wesentlichen identisch zur Rückfaltungsreaktion in Anwesenheit von DTT (sieheAbb. 83 A). Aufgrund des kleineren Reduktionspotentials für die Rückfaltung inklusive TCEP erfolgt die Bildung des rhM-CSF Dimers etwas schneller. Bereits nach ca. 1 h liegt das rhM-CSF zur Hälfte in Form des Dimeren vor. Dagegen zeigt die SDS-PAGE (nicht reduzierend) der mit MEL modifizierten Faltungsintermediate für die Zeitpunkte 0 h bis 42 h einen höheren Monomer- und dementsprechend geringeren Dimeranteil. Auch hier erfolgte durch die SDS-PAGE-Probenpräparation (siehe 3.6.4) eine partielle Re-reduktion bereits zurückgefalteter rhM-CSF Monomere und Dimere durch das noch in der Reaktionslösung vorliegende, nicht modifizierte GSH (s.o.).
Das durchschnittliche Molekulargewicht der modifizierten rhM-CSF Faltungsintermediate wurde analog der Rückfaltung mit DTT-reduziertem rhM-CSF berechnet (siehe S. 93), und daraus die durchschnittlichen Modifizierungsgrade bzw. die Anzahl der bereits ausgebildeten Disulfidbrücken für die IAA-Alkylierung (Tab. 10) und für die MEL-Modifizierung (Tab. 11) abgeleitet.

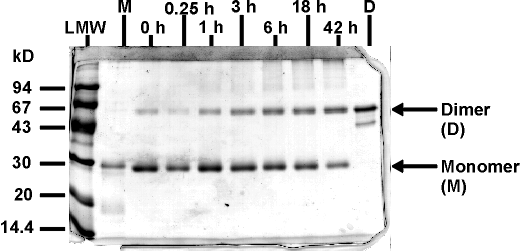
Abb. 86: SDS-PAGE (12% Lämmli) unter nicht reduzierenden Bedingungen der Rückfaltungsreaktion von TCEP reduziertem rhM-CSF. Die Faltungsintermediate nach 0 h bis 42 h wurden durch (A)IAA-Alkylierung (B)MEL-Modifizierung stabilisiert. Aufgetragen sind die Reaktionslösungen vor der Ultrafiltration (entsprechend 1 µg). Mit LMW ist der Molekulargewichtsmarker gekennzeichnet. M entspricht dem reduzierten, aber nicht modifizierten rhM-CSF Monomer (2 µg); D dem intakten rhM-CSF Dimer (2 µg).
Tab. 10: MALDI-MS Molekulargewichtsbestimmungen der carboxamidomethylierten Faltungsintermediate des mit TCEP reduzierten rhM-CSF (nach Ultrafiltration). Die m/z-Werte geben das durchschnittlich Molekulargewicht an (siehe Text). Die Werte für n(IAA) bzw. m(SS) geben die Anzahl der eingeführten Carboxamidomethylierungen bzw. der bereits ausgebildeten Disulfidbrücken bezogen auf das Dimer an.
|
[DH2]2+ / [MH]+ |
[DH]+ |
|||||
|
zeitlicher |
m/z(Durchschn.) |
n(IAA) bez. auf Dimer |
m(SS) bez. auf Dimer |
m/z (Durchschn.) |
n(IAA) bez. auf Dimer |
m(SS) bez. auf Dimer |
|
0 h |
25054 |
18.6 |
0 |
- |
- |
- |
|
0.25 h |
24911 |
13.6 |
2-3 |
- |
- |
- |
|
1 h |
24676 |
5.6 |
6-7 |
49355 |
5.6 |
6-7 |
|
3 h |
24549 |
1.2 |
8-9 |
49197 |
2.9 |
7-8 |
|
6 h |
24571 |
2.0 |
8 |
49110 |
1.4 |
8-9 |
|
18 h |
24577 |
2.2 |
8 |
49141 |
1.9 |
8 |
|
42 h |
24573 |
2.0 |
8 |
49155 |
2.2 |
8 |
Tab. 11: MALDI-MS Molekulargewichtsbestimmungen der MEL-modifizierten Faltungsintermediate pro Modifizierungsgrad (siehe Text) des mit TCEP reduzierten rhM-CSF (nach Ultrafiltration). Die Werte für n(MEL) bzw. m(SS) geben die Anzahl der eingeführten MEL-Modifizierungen bzw. der bereits ausgebildeten Disulfidbrücken bezogen auf das Dimer an.
|
[DH2]2+ / [MH]+ |
[DH]+ |
|||||
|
zeitlicher Verlauf |
m/z(Durchschn.) |
n(MEL) bez. auf Dimer |
m(SS) bez. auf Dimer |
m/z (Durchschn.) |
n(MEL) bez. auf Dimer |
m(SS) bez. auf Dimer |
|
0 h |
25653 |
8.2 |
0-1 |
- |
- |
- |
|
25368 |
6.2 |
2-3 |
||||
|
0.25 h |
25649 |
8.2 |
0-1 |
- |
- |
- |
|
25371 |
6.2 |
2-3 |
||||
|
1 h |
25391 |
6.4 |
2-3 |
49948 |
3.5 |
5-6 |
|
25124 |
4.4 |
4-5 |
||||
|
24828 |
2.2 |
6-7 |
||||
|
3 h |
25114 |
4.4 |
4-5 |
49953 |
3.3 |
5-6 |
|
24869 |
2.6 |
6-7 |
||||
|
6 h |
25109 |
4.4 |
4-5 |
49344 |
1.1 |
8 |
|
24853 |
2.4 |
6-7 |
||||
|
18 h |
24835 |
2.4 |
6-7 |
49410 |
1.4 |
7-8 |
|
24523 |
0 |
9 |
49092 |
0.2 |
8-9 |
|
|
42 h |
24838 |
2.4 |
6-7 |
49123 |
0.34 |
8-9 |
|
24632 |
0.8 |
7-8 |
||||
Die aus der MALDI-MS-Analyse bestimmten durchschnittlich eingeführten Carboxamidomethylierungen (Tab. 10) geben die schnellere Rückfaltung des rhM-CSF gut wieder. So liegen nach ca. 1 h durch sechs bzw. sieben der neun Disulfidbrücken des rhM-CSF Dimers geschlossen vor (vgl. mit Tab. 8). Die nach 6 h bis 42 h gefundene zweifache Carboxamidomethylierung, bezogen auf das rhM-CSF Dimer, könnte durch die Carboxamidomethylierung der zwei N-Termini des Homodimers erklärt werden. Auch die Anzahl der durchschnittlich eingeführten MEL-Modifizierungen (Tab. 11) geben den schnelleren Verlauf der Rückfaltung wieder (vgl. mit Tab. 9).
Für die mit TCEP reduziertem rhM-CSF durchgeführte Rückfaltung erhält man für alle Zeitpunkte der Rückfaltungskinetik eine größere Anzahl an MEL-Modifizierungen als Carboxamidomethylierungen. Die aus den durchschnittlichen Modifizierungsgraden berechnete Anzahl bereits ausgebildeter Disulfidbrücken ist für die mit MEL modifizierten Faltungsintermediate durchschnittlich kleiner (sieheTab. 10 und Tab. 11). Vermutlich erfolgt hier die partielle Re-reduktion bereits ausgebildeter Disulfidbrücken der mit MEL-modifizierten Faltungsintermediate durch GSH, da MEL keine stabilen Arsonigsäuredithioester mit GSH bildet (siehe S. 87). Diese partielle Re-reduktion kann nur unter basischen Bedingungen erfolgen, da nur dann das GSH-Thiolatanion vorliegt. Die erneut gebildeten Proteinbisthiole reagieren dann mit noch vorhandenem MEL. Daraus folgt, daß diese zusätzlichen MEL-Modifizierungen keine Artefakte der Probenaufarbeitung, also Ultrafiltration und Umpuffern in ein Lösungsmittelgemisch mit pH 2 8 (s.o.), sind, sondern während der Modifizierungsreaktion eintreten müssen. Dies wird durch die vergleichbaren MALDI-Massenspektren der MEL-modifizierten Faltungsintermediate vor und nach Ultrafiltration bestätigt. Daß dieser Effekt nicht bei der mit DTT reduziertem rhM-CSF durchgeführten Rückfaltung zu beobachten ist, beruht auf der quantitativen Ausbildung des zyklischen MEL-DTT-Addukts.
Da MEL sehr schnell mit Biscysteinylfunktionen in Peptiden (siehe 2.2.3) und Proteinen (siehe 2.6) reagiert, könnte diese Artefaktbildung durch eine kürzere Reaktionszeit zwischen MEL und den Faltungsintermediaten und anschließendes Ansäuern der Reaktionslösung vermieden werden. Eine weitere Möglichkeit diese Re-reduktion bereits gebildeter Disulfidbrücken zu vermeiden, wäre die Alkylierung des freien Monothiols GSH, nach der erfolgten MEL-Modifizierung der Faltungsintermediate. Wie in Abschnitt 2.5 gezeigt, kann MEL als Schutzgruppe für Biscysteinylfunktionen in Peptiden gegen Alkylierungsmittel fungieren.
|
|
|
|
|
2.8 Computerberechnungen von lösungsmittelzugänglichen Oberflächen in Proteinen mit bekannter Struktur |
|
|
2.6 Selektive chemische Modifizierung von Modellproteinen mit Melarsenoxid und massenspektrometrische Charakterisierung der Reaktionsprodukte |
Diese Seite wurde erstellt am 15. März 1998. Letzte Änderungen am 30. Juni 1998 durchPeter Happersberger.