|
|
|
|
|
2.3 Massenspektrometrische Charakterisierung der Reaktionsprodukte von Modellpeptiden mit Arsonigen Säuren |
|
|
2.1 Synthese von Derivaten der Arsonigen Säure |
2.2 UV-spektroskopische Untersuchungen zum Reaktionsmechanismus der Modifizierung von Bis-Thiolen mit Melarsen Oxid
Wie in Abschnitt 1.2.3 erwähnt reagieren Arsonige Säuren mit Dithiolen zu zyklischen Arsonigsäuredithioester, wobei die entstehende Ringgröße vom eingesetzten Dithiol abhängt. Systematische Untersuchungen in der Literatur beschränken sich meist auf die Verwendung von 1,2-Dithiolen [69], 1,3-Dithiolen [60] und 1,4-Dithiolen [127]. Dabei bestätigt sich insbesondere die bevorzugte Stabilität des Fünfringes. Die Ausbildung des Siebenrings (1,4-Dithiole) scheint sterisch ungünstig zu sein [59, 127]. Untersuchungen von V. P. Whittaker (1947) mit ,-Dithiolen, die zu Ringgrößen von 5-15 Atomen führten, zeigen wieder eine erhöhte Stabilität ab dem Achtring, die aber nicht an die des Fünfrings heranreicht, und tendenziell mit zunehmender Ringgröße abnimmt. Bei der biscysteinylselektiven Modifizierung von Proteinen mit Arsonigen Säuren können jedoch weit größere Makrozyklen resultieren. So erhält man z.B. für das Peptid Oxytocin (Abb. 34) mit zwei Cysteinen, die durch vier weitere Aminosäuren voneinander getrennt sind, einen Makrozyklus aus 21 Atomen. Für jede weiter dazukommende Aminosäure zwischen zwei Cysteinen vergrößert sich der Makrozyklus um drei weitere Atome. Analogiebetrachtungen zu den Carbozyklen zeigen, daß dort die höchste Stabilität für den Fünf- und Sechsring auftritt. Mittlere Ringgrößen (8-12 Atome) zeigen aufgrund sterischer Wechselwirkungen geringere Stabilitäten, während Makrozyklen (> 12 Ringglieder), z.B. Kronenether, durchaus sehr stabil sind, da sich die C-Atome unter Beibehaltung des normalen Tetraederwinkels in zwei fast parallelen C-Zickzack-Ketten anordnen können [128]. Für Ringe, die Heteroatome enthalten gelten diese Regeln nur eingeschränkt. C-Ringe, die zusätzlich nur Arsen und Schwefel enthalten sind wegen den fehlenden H-Atomen vermutlich stabiler [59]. In Peptiden und Proteinen ist der Sachverhalt durch (fast) planare Peptidbindungen nochmals komplizierter. Die Isolierung des Melarsen Oxid-Trypanothion-Addukts (siehe S. 11) zeigt, daß Makrozyklen aus Peptiden mit 25 Atomen durchaus stabil sind [69].
Folgende, für die spätere Charakterisierung der Proteinfaltungsreaktion relevante Fragestellungen sollten mit der UV-spektroskopischen Untersuchung der Reaktion von Melarsen Oxid (MEL) mit Dithiolen untersucht werden:
- Beeinflußt die entstehende Ringgröße des zyklischen Arsonigsäuredithioesters Reaktionsgeschwindigkeit und Stöchiometrie der Reaktion?
- Sind Aussagen über die Stöchiometrie und den zeitlichen Verlauf der Reaktion in Abhängigkeit des pH-Werts möglich?
Als Dithiole sollten verwendet werden: 2,3-Dimercaptopropanol (DMP) und 1,4-Dithiothreitol bzw. nach Reduktion das Peptid Oxytocin (sieheAbb. 34).

Abb. 34: Strukturformeln der Dithiole 2,3-Dimercaptopropanol (DMP) und 1,4-Dithiothreitol (DTT). Schematisch ist die Struktur des zyklischen Peptids Oxytocin (OT) gezeigt.
2.2.1 Die Cystinreduktionsmittel DTT und TCEP
Oxytocin, ein aus neun Aminosäuren bestehendes Peptid, besitzt eine zyklische Struktur, die durch eine Disulfidbrücke stabilisiert wird (siehe Abb. 34). Erst nach Reduktion liegt ein Dithiol vor. Analog mußten Vasopressin (siehe 2.3.2), sowie die verwendeten Modellproteine (siehe 2.6.2) vor der Reaktion mit den hier synthetisierten Derivaten der Arsonigen Säure reduziert werden. Typische Reduktionsmittel sind selbst Thiole, wie 1,4-Dithiothreitol (DTT) oder 2-Mercaptoethanol (ME) [129,130]. Der Thiol-Disulfid Austausch - dargestellt in Abb. 35 für DTT - erfolgt in zwei Schritten. Entscheidend für die Reduktion ist die Bildung des Thiolatanions, da dieses im ersten Schritt mit der Cystinylgruppe des Proteins zu einem gemischten Disulfid reagiert. Dieses reagiert dann unter Ausbildung des thermodynamisch günstigen Sechsringes von oxidiertem DTT zur Biscysteinylfunktion am Protein. Aus dem Vergleich der pKa-Werte von DTT (pKa1 = 8.3; pKa2 = 9.5) mit ME (pKa = 9.5) und dem negativeren Redoxpotential von DTT folgt, daß DTT das effektivere Reduktionsmittel ist [130].
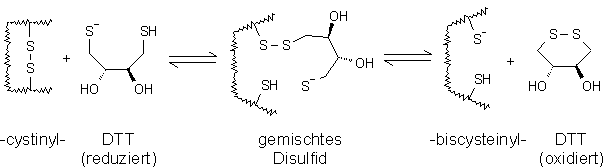
Abb. 35: Reaktionsschema der Reduktion von Cystinylgruppen in Proteinen durch 1,4-Dithiothreitol (DTT) zu Biscysteinylgruppen.
Die Verschiebung des Gleichgewichts auf die Seite des reduzierten Proteins erfolgt für DTT bereits bei geringen molaren Überschüssen (ungefähr 10-20-fach pro Disulfidbrücke). Im Gegensatz dazu muß bei der Reduktion mit 2-Mercaptoethanol ein hoher molarer Überschuß (100-500 fach) eingesetzt werden, um das Gleichgewicht auf die Seite des reduzierten Proteins zu verschieben [130]. Beide Reduktionsmittel arbeiten nur für pH-Werte größer sieben effektiv. Ein weiterer Nachteil der Thiolreduktionsmittel ist ihre Reaktivität gegenüber nachfolgenden cysteinylselektiven proteinchemischen Umsetzungen, z.B. reagiert DTT mit Arsonigen Säuren (siehe 2.3.2), so daß diese in deutlich höheren molaren Verhältnissen zugesetzt werden müssen. Damit wäre die UV-spektroskopische Verfolgung des Reaktionsverlauf erschwert. Für viele Alkylierungsmittel (z.B. 4-Vinylpyridin), kann gerade dieser größere molare Überschuß zu unerwünschten Nebenreaktionen führen, Alkylierungen weiterer nukleophiler Gruppen im Protein sind möglich [131].
Ein Cystinylreduktionsmittel, das nicht auf der Ausbildung gemischter Thiol-Disulfidstrukturen beruht ist das wasserlösliche Trialkylphosphin Tris-(2-carboxyethyl)phosphin (TCEP) [132,133]. TCEP reduziert selektiv Disulfidbrücken (siehe Abb. 36) bei deutlich geringeren molaren Verhältnissen als für DTT.
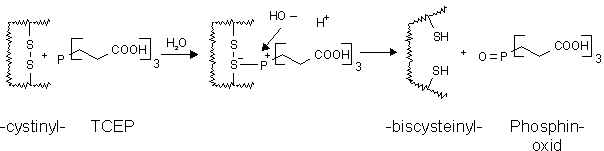
Abb. 36: Reaktionsschema der Reduktion von Cystinylgruppen in Proteinen durch Tris-(2-carboxyethyl)phosphin (TCEP) zu Biscysteinylgruppen [131].
Dabei reagiert TCEP irreversibel zum thermodynamisch stabileren Phosphinoxid. Die Reduktion erfolgt schneller, effektiver und auch noch bei pH-Wertenkleiner Sieben [134]. TCEP reagiert nicht mit dem Alkylierungsreagenz 4-Vinylpyridin, reagiert aber mit Iodessigsäure zu einem quaternären Phosphonium-Salz [131]. Bestimmt man das Differenzspektrum (siehe 2.2.2) von TCEP und MEL so erhält man eine (fast) waagrechte Kurve, d.h. TCEP und MEL reagieren nicht miteinander. Somit ermöglicht TCEP - ohne vorherige Abtrennung - die UV-spektroskopische Analyse des Reaktionsverlaufs der Modifizierung von reduzierten Proteinen mit Arsonigen Säuren.
2.2.2 Analyse des Reaktionsverlaufs - Chemische Selektivität und Stöchiometrie der Modifizierungsreaktion mit Melarsen Oxid
Für die UV-spektroskopische Untersuchung der Reaktion von Dithiolen mit Melarsen Oxid (MEL) wurden die niedermolekularen Dithiole 2,3-Dimercaptopropanol (DMP; British Anti Lewisite BAL), 1,4-Dithiothreitol (DTT) und das Peptid Oxytocin (OT) ausgesucht (sieheAbb. 34). Bei der Ausbildung der Arsonigsäuredithioester mit MEL verschiebt sich das Absorptionsmaximum von ![]() max = 273 nm für MEL auf ungefähr
max = 273 nm für MEL auf ungefähr ![]() max = 280 nm für das MEL-Dithioladdukt. Dieser Wert variiert leicht in Abhängigkeit des verwendeten Dithiols und pH-Werts (277-281 nm) und stimmt mit dem in der Literatur genannten
max = 280 nm für das MEL-Dithioladdukt. Dieser Wert variiert leicht in Abhängigkeit des verwendeten Dithiols und pH-Werts (277-281 nm) und stimmt mit dem in der Literatur genannten ![]() max = 282 nm gut überein [69]. Das angewendete Verfahren am Beispiel der Umsetzung von OT mit MEL wird beschrieben. Die Reduktion von OT erfolgte durch TCEP.
max = 282 nm gut überein [69]. Das angewendete Verfahren am Beispiel der Umsetzung von OT mit MEL wird beschrieben. Die Reduktion von OT erfolgte durch TCEP.
Melarsen Oxid (MEL) (je 45 µM) wurde mit verschiedenen molaren Verhältnissen an Oxytocin (OT) in wäßrigen Puffersystemen umgesetzt und der Reaktionsverlauf UV-spektroskopisch verfolgt. Die Konzentration von OT wurde über die OT-Einwaage bestimmt. Es wurden UV-Spektren im Bereich von ![]() = 201-339 für verschiedene Konzentrationen n an OT (Standard OTn) und für die konstant gehaltene Konzentration von MEL (Standard M, 45 µM) separat aufgenommen. Für die verschiedenen molaren Verhältnisse bildet manDifferenzspektren (Gl.1) aus den UV-Spektren des MEL-haltigen Produkts (Produktgemisch Pn) und den Summenspektren der Einzelkomponenten (siehe Abb. 37).
= 201-339 für verschiedene Konzentrationen n an OT (Standard OTn) und für die konstant gehaltene Konzentration von MEL (Standard M, 45 µM) separat aufgenommen. Für die verschiedenen molaren Verhältnisse bildet manDifferenzspektren (Gl.1) aus den UV-Spektren des MEL-haltigen Produkts (Produktgemisch Pn) und den Summenspektren der Einzelkomponenten (siehe Abb. 37).
Differenzspektrum = Produktgemisch Pn - (Standard M + Standard OTn) (Gl. 1)
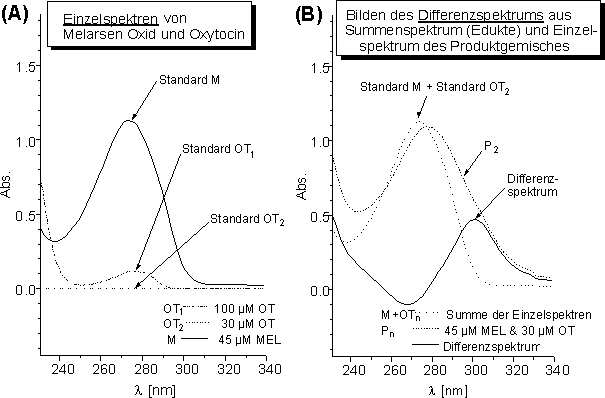
Abb. 37: (A) Einzelspektren von 45 µM MEL (Standard M) und 30 bzw. 100 µM OT (Standard OTn) in 10 mM NH4HCO3 pH 8.2 (ctcep = 150 µM). (B) Bilden des Differenzspektrums aus der Summe der Einzelspektren (Standard M und Standard OT2) und dem Produktgemisch von MEL und OT.
Überlagert man die Differenzspektren für verschiedene molare Verhältnisse von MEL und OT in einem Diagramm ergeben sich Kreuzungsbereiche (KB) mit zugehörigem Minimum und Maximum (Abb. 38 A). Die Lagen der Minima (![]() = 264-268 nm) und Maxima (
= 264-268 nm) und Maxima (![]() = 301-303 nm) verschieben sich in Abhängigkeit des verwendeten Dithiols und dem pH-Wert des verwendeten Puffersystems leicht. Aus der Differenz der Absorptionen für das Minimum bei 268 nm und Maximum bei 301 nm (hier pH 8.2) wird der Anteil des gebildeten MEL-OT-Produkts abgeleitet. Diese Differenz wird alsAbsorptionsdifferenz OD bezeichnet (Gl. 2).
= 301-303 nm) verschieben sich in Abhängigkeit des verwendeten Dithiols und dem pH-Wert des verwendeten Puffersystems leicht. Aus der Differenz der Absorptionen für das Minimum bei 268 nm und Maximum bei 301 nm (hier pH 8.2) wird der Anteil des gebildeten MEL-OT-Produkts abgeleitet. Diese Differenz wird alsAbsorptionsdifferenz OD bezeichnet (Gl. 2).
![]() OD301-268 = A 301 nm - A 268 nm (Gl. 2)
OD301-268 = A 301 nm - A 268 nm (Gl. 2)
Mit zunehmender Dithiolkonzentration, also Oxytocinkonzentration, nimmt der Anteil des MEL-OT-Produkts, d.h. ![]() OD301-268 zu. Bei der vollständigen Umsetzung von MEL erhält man den maximalen Betrag von
OD301-268 zu. Bei der vollständigen Umsetzung von MEL erhält man den maximalen Betrag von ![]() OD301-268 (siehe Abb. 38). Um die erhaltenen Ergebnisse mit der Literatur und mit Peptiden/Proteinen mit mehr als zwei Thiolfunktionen zu vergleichen werden die so ermittelten
OD301-268 (siehe Abb. 38). Um die erhaltenen Ergebnisse mit der Literatur und mit Peptiden/Proteinen mit mehr als zwei Thiolfunktionen zu vergleichen werden die so ermittelten ![]() OD301-268-Werte in Abhängigkeit der Thiolkonzentration (= der doppelten Oxytocinkonzentration) aufgetragen.Abb. 38 B zeigt einen linearen Anstieg der Kurve, solange die Thiolkonzentration die MEL-Konzentration nicht übersteigt. D.h. MEL reagiert proportional zur eingesetzten Thiolkonzentration zum Arsonigsäuredithioester. Die berechnete Gerade für diesen Teil der Funktion bestätigt den theoretisch notwendigen Nulldurchgang.
OD301-268-Werte in Abhängigkeit der Thiolkonzentration (= der doppelten Oxytocinkonzentration) aufgetragen.Abb. 38 B zeigt einen linearen Anstieg der Kurve, solange die Thiolkonzentration die MEL-Konzentration nicht übersteigt. D.h. MEL reagiert proportional zur eingesetzten Thiolkonzentration zum Arsonigsäuredithioester. Die berechnete Gerade für diesen Teil der Funktion bestätigt den theoretisch notwendigen Nulldurchgang.

Abb. 38: (A) Differenzspektrenschar der Umsetzung von OT mit MEL für verschiedene molare Verhältnisse (siehe Legende) im NH4HCO3-Puffer bei pH 8.2. Die -Werte der Minima/Maxima der Differenzspektren sind identisch. Die Differenz der Absorption des Minimums (268 nm) und des Maximums (301 nm) ![]() OD301-268 nm ist für das molare Verhältnis (MEL / OT 45:45 ) dargestellt. (B) Dargestellt sind die entsprechenden
OD301-268 nm ist für das molare Verhältnis (MEL / OT 45:45 ) dargestellt. (B) Dargestellt sind die entsprechenden ![]() OD301-268 nm-Werte für die gesamte Differenzspektrenschar in (A) in Abhängigkeit der Thiolkonzentration (doppelte Dithiolkonzentration).
OD301-268 nm-Werte für die gesamte Differenzspektrenschar in (A) in Abhängigkeit der Thiolkonzentration (doppelte Dithiolkonzentration).
Überschreitet die Thiolkonzentration die eingesetzte MEL- Konzentration, so erfolgt die Umsetzung so lange, bis MEL vollständig als MEL-OT-Produkt vorliegt, d.h. ![]() OD301-268 steigt nicht mehr, die Kurve verflacht in ein Plateau. Die Kurve knickt bei der doppelten MEL-Konzentration ab und zeigt, daß die Umsetzung von Melarsen Oxid mit Oxytocin zu zyklischen Arsonigsäuredithioester in einer 1:1 Stöchiometrie erfolgt.
OD301-268 steigt nicht mehr, die Kurve verflacht in ein Plateau. Die Kurve knickt bei der doppelten MEL-Konzentration ab und zeigt, daß die Umsetzung von Melarsen Oxid mit Oxytocin zu zyklischen Arsonigsäuredithioester in einer 1:1 Stöchiometrie erfolgt.
Es sollte nun untersucht werden, welchen Einfluß der pH-Wert auf die 1:1 Stöchiometrie der Reaktion von MEL mit OT hat. Melarsen Oxid wurde mit unterschiedlichen molaren Verhältnissen an TCEP reduziertem Oxytocin bei folgenden pH-Werten umgesetzt: 10 mM NH4HCO3pH 8.2, 10 mM NH4OAc pH 6.0 und 4.0 und 10 mM Glycin pH 2.0 . Die Reaktion wurde UV-spektroskopisch verfolgt und die ![]() ODmax-min Werte analog dem oben beschriebenen Verfahren bestimmt. Die entsprechende Verschiebung von Minimum und Maximum bei verschiedenen pH-Werten wurde berücksichtigt (siehe Legende inAbb. 39 A). Die Abhängigkeit von
ODmax-min Werte analog dem oben beschriebenen Verfahren bestimmt. Die entsprechende Verschiebung von Minimum und Maximum bei verschiedenen pH-Werten wurde berücksichtigt (siehe Legende inAbb. 39 A). Die Abhängigkeit von ![]() ODmax-min von der freien Thiolkonzentration für die Reaktion von MEL mit Oxytocin bei verschiedenen pH-Werten zeigtAbb. 39 A. Zum Vergleich wurde
ODmax-min von der freien Thiolkonzentration für die Reaktion von MEL mit Oxytocin bei verschiedenen pH-Werten zeigtAbb. 39 A. Zum Vergleich wurde ![]() ODmax-min für die Reaktion von MEL mit demMonothiol Glutathion (L--Glutamyl-L-cysteinyl-glycin; GSH) bei pH 8.5 bestimmt (Abb. 39 B).
ODmax-min für die Reaktion von MEL mit demMonothiol Glutathion (L--Glutamyl-L-cysteinyl-glycin; GSH) bei pH 8.5 bestimmt (Abb. 39 B).
Unabhängig vom jeweiligen pH-Wert steigen die Kurven in guter Näherung linear mit zunehmender Thiolkonzentration an Oxytocin. Da die Reaktion von MEL mit OT bei pH 2 deutlich langsamer als für höhere pH-Werte verläuft (siehe 2.2.3), wird das Plateau der Kurve erst nach längerer Reaktionszeit erreicht. Das Abknicken der Kurven erfolgt für alle pH-Werte bei der Thiolkonzentration von ca. 90 µM, also ungefähr der doppelten MEL-Konzentration (45 µM). D.h. die Umsetzung von Arsonigen Säuren mit Dithiolen zu zyklischen Arsonigsäuredithioester erfolgt - pH-unabhängig - in einer 1:1 Stöchiometrie. Identische Untersuchungen wurden für die Reaktion von MEL mit den niedermolekularen Dithiolen DMP und DTT durchgeführt (siehe 3.3.2). Die gefundenen Ergebnisse verhalten sich analog zu OT und stimmen mit der Literatur überein [69]. Unabhängig von der entstehenden Ringgröße des Reaktionsproduktes erhält man eine 1:1 Stöchiometrie für MEL und dem jeweiligen Dithiol, d.h. die vollständige Umsetzung von MEL erfolgt bereits für sehr geringe molare Überschüsse an Dithiol. Dagegen erfolgt die vollständige Umsetzung von GSH mit MEL (Abb. 39 B) erst bei sehr viel höheren Thiolkonzentrationen (ca. 500 µM, Daten nicht gezeigt) und nähert sich nur langsam der maximalen Absorptionsdifferenz ![]() OD299-267. Dies bestätigt die geringere Stabilität der offenkettigen Arsonigsäuredithioester. Auch die Ausbildung makrozyklischer Arsonigsäuredithioester ist den offenkettigen bevorzugt, denn gibt man zu einer stöchiometrischen 1:1 Mischung von MEL und GSH reduziertes Oxytocin, so erfolgt sofort quantitativ die Ausbildung des zyklischen Oxytocin-Arsonigsäurediesters (siehe 2.7.3).
OD299-267. Dies bestätigt die geringere Stabilität der offenkettigen Arsonigsäuredithioester. Auch die Ausbildung makrozyklischer Arsonigsäuredithioester ist den offenkettigen bevorzugt, denn gibt man zu einer stöchiometrischen 1:1 Mischung von MEL und GSH reduziertes Oxytocin, so erfolgt sofort quantitativ die Ausbildung des zyklischen Oxytocin-Arsonigsäurediesters (siehe 2.7.3).
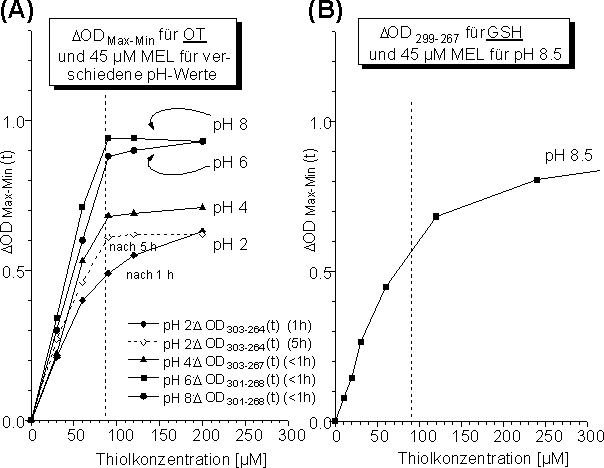
Abb. 39: (A) Umsetzungsgrad von MEL mit OT in Abhängigkeit der Thiolkonzentration für verschiedene pH-Werte. In der Legende sind die Wellenlängen des Maximums und Minimums des Differenzspektrums beim jeweiligen pH-Wert angegeben; die Zeitangabe entspricht der Reaktionszeit. (B) Umsetzungsgrad von Melarsen Oxid mit dem Monothiol Glutathion (GSH) bei pH 8.5.
2.2.3 Zeitlicher Verlauf der Reaktion in Abhängigkeit des pH-Werts und des verwendeten Puffersystems
Um den zeitlichen Verlauf der Reaktion von Melarsen Oxid (MEL) mit den Dithiolen UV-spektroskopisch zu verfolgen, mußte die zeitliche Veränderung von ![]() ODmax-min, also
ODmax-min, also ![]() ODmax-min(t) bestimmt werden. Aus praktischen Gründen wurde die zeitliche Veränderung der Absorptionswerte nur bei 267 nm und 299 nm und nicht für das komplette Spektrum bestimmt. Aus den so gewonnenen Daten kann
ODmax-min(t) bestimmt werden. Aus praktischen Gründen wurde die zeitliche Veränderung der Absorptionswerte nur bei 267 nm und 299 nm und nicht für das komplette Spektrum bestimmt. Aus den so gewonnenen Daten kann ![]() OD299-267(t) berechnet werden.
OD299-267(t) berechnet werden.
In Abb. 40 ist das Vorgehen am Beispiel MEL und Oxytocin (OT) für die Umsetzung im NH4HCO3-Puffer bei pH 8.2 gezeigt. Eine 45 µM MEL-Lösung wurde in der UV-Küvette vorgelegt und die simultane Messung der Absorptionswerte A267 nm(t) und A299 nm(t) gestartet (Abb. 40 A, t < 50 s). Für t = 50 s wurde 30 µM reduziertes (TCEP) Oxytocin zugegeben. Da dazu das UV-Photometer geöffnet werden mußte, resultiert ein Zeitfenster (gestrichelte Linien in Abb. 40 A) von ungefähr 20 Sekunden, in dem keine Absorptionswerte bestimmt werden konnten. Man erkennt deutlich, daß die Reaktion von MEL und OT bei pH 8.2 innerhalb von 20 s quantitativ erfolgt, da sich die Absorptionswerte A267 nm(t) und A299 nm(t) praktisch nicht mehr ändern. Die aus diesen Werten berechnete zeitliche Änderung von ![]() OD299-267(t) ist in Abb. 40 B (Kurve 2) dargestellt.
OD299-267(t) ist in Abb. 40 B (Kurve 2) dargestellt.
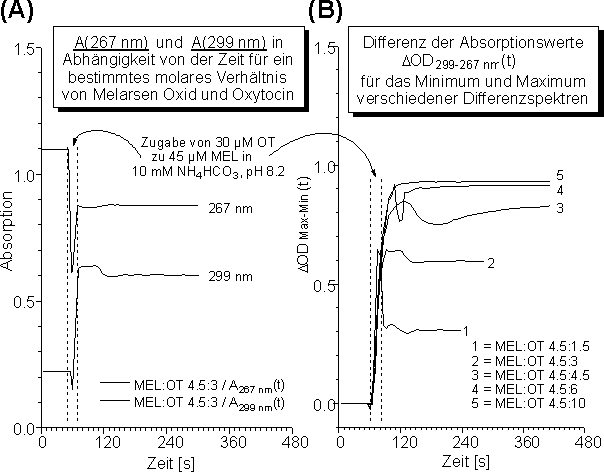
Abb. 40: (A) Zeitlicher Verlauf der Absorption bei 267 und 299 nm für die Umsetzung von MEL mit OT im molaren Verhältnis 45:30 in 10 mM NH4HCO3 pH 8.2. Die gestrichelten vertikalen Linien bei t 50 s geben den Zeitraum von ca. 20 s zum Öffnen des UV-Photometers und zur Zugabe des Dithiols an. (B) Zeitlicher Verlauf von ![]() OD299-267 (t) für die in der Legende angegebenen molaren Verhältnisse von MEL und OT in 10 mM NH4HCO3 pH 8.2.
OD299-267 (t) für die in der Legende angegebenen molaren Verhältnisse von MEL und OT in 10 mM NH4HCO3 pH 8.2.
Ferner ist in Abb. 40 B die zeitliche Änderung von ![]() OD299-267(t) für verschiedene molare Verhältnisse von MEL zu OT (siehe Legende) gezeigt. Für molare Verhältnisse größer der 1:1 Stöchiometrie erhält man einen maximalen Wert für
OD299-267(t) für verschiedene molare Verhältnisse von MEL zu OT (siehe Legende) gezeigt. Für molare Verhältnisse größer der 1:1 Stöchiometrie erhält man einen maximalen Wert für ![]() OD299-267(t) (s.o.) Am Ende der Reaktion wurde zur Kontrolle das Gesamtdifferenzspektrum ( = 201-339 nm) aufgenommen und daraus unabhängig erneut
OD299-267(t) (s.o.) Am Ende der Reaktion wurde zur Kontrolle das Gesamtdifferenzspektrum ( = 201-339 nm) aufgenommen und daraus unabhängig erneut ![]() OD299-267 bestimmt. Die erhaltenen Werte waren in guter Übereinstimmung (OD299-267 0.02).
OD299-267 bestimmt. Die erhaltenen Werte waren in guter Übereinstimmung (OD299-267 0.02).
Der zeitliche Verlauf der Reaktion von MEL mit Oxytocin wurde für verschiedene molare Verhältnisse pH-abhängig bestimmt. Analog der oben beschriebenen Methode wurde eine konstant gehaltene Menge an MEL (je 45 µM) bei folgenden pH-Werten mit verschiedenen Konzentrationen an reduziertem (TCEP) Oxytocin umgesetzt: pH 8.2, pH 6.0,4.0 und pH 2.0. Es wurden sowohl die oben beschriebenen Puffersysteme als auch NaH2PO4-Puffer verwendet. Der Reaktionsverlauf wurde UV-spektroskopisch verfolgt und ![]() OD299-267(t) bestimmt. Unabhängig vom verwendeten Puffersystem erhält man vergleichbare Ergebnisse.Abb. 41 zeigt den ph-abhängigen zeitlichen Verlauf der Reaktion von MEL mit Oxytocin. Die Umsetzung von MEL mit Oxytocin erfolgt bei pH 8 und pH 6 innerhalb von maximal 20 Sekunden (Zeitfenster zum Öffnen des UV-Photometers) quantitativ. Mit abnehmendem pH-Wert verlangsamt sich die Reaktion deutlich. Bei pH 2 erfolgt selbst nach einer Stunde Reaktionszeit keine quantitative Umsetzung.
OD299-267(t) bestimmt. Unabhängig vom verwendeten Puffersystem erhält man vergleichbare Ergebnisse.Abb. 41 zeigt den ph-abhängigen zeitlichen Verlauf der Reaktion von MEL mit Oxytocin. Die Umsetzung von MEL mit Oxytocin erfolgt bei pH 8 und pH 6 innerhalb von maximal 20 Sekunden (Zeitfenster zum Öffnen des UV-Photometers) quantitativ. Mit abnehmendem pH-Wert verlangsamt sich die Reaktion deutlich. Bei pH 2 erfolgt selbst nach einer Stunde Reaktionszeit keine quantitative Umsetzung.
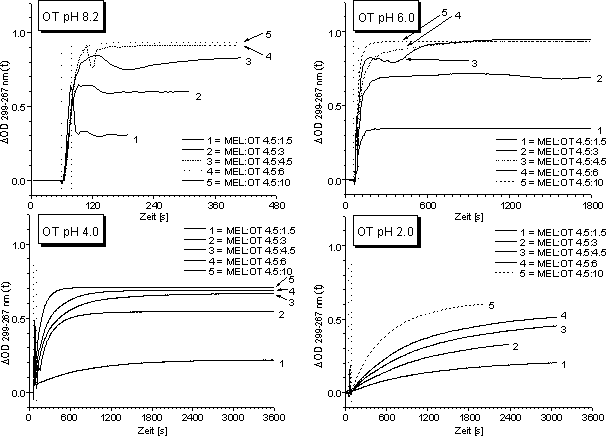
Abb. 41: Zeitlicher Verlauf der Reaktion von 45 µM MEL mit verschiedenen Konzentration an OT für verschiedene pH-Werte. Die Legende gibt die molaren Verhältnisse wieder. Die gestrichelten vertikalen Linien entsprechen der Zugabe des Dithiols zur MEL-Lösung (Öffnen des UV-Photometers).

Abb. 42: Zeitlicher Verlauf der Reaktion von 45 µM MEL mit verschiedenen Konzentration an DTT für verschiedene pH-Werte. Die Legende gibt die molaren Verhältnisse wieder. Die gestrichelten vertikalen Linien entsprechen der Zugabe des Dithiols zur MEL-Lösung (Öffnen des UV-Photometers).
Analoge Untersuchungen wurden für die niedermolekularen Dithiole DMP und DTT durchgeführt, deren Ergebnisse den hier beschriebenen entsprechen. Stellvertretend zeigt Abb. 42 die Absorptionsdifferenz ![]() OD299-267(t) für die Reaktion von MEL mit DTT (für DMP: siehe 3.3.2). Auch hier erfolgt die Umsetzung bei pH 8 innerhalb von 20 Sekunden (Zeitfenster). Bei pH 6 und ausgeprägter bei pH 4 nimmt die Reaktionsgeschwindigkeit deutlich ab, bei pH 2 jedoch wieder zu. Die Zunahme der Reaktionsgeschwindigkeit der Reaktion von Melarsen Oxid mit den niedermolekularen Dithiolen bei niedrigen pH-Werten kann durch eine säurekatalysierte Veresterung - analog der Bildung von Carbonsäureestern - beschrieben werden. Mit zunehmenden pH-Wert würde sich dann die Geschwindigkeit der Reaktion verlangsamen. Die sehr viel größere Reaktionsgeschwindigkeit im Basischen läßt sich auf die deutlich höhere Nukleophilie des Thiolatanions (pKa 7-9) zurückführen.
OD299-267(t) für die Reaktion von MEL mit DTT (für DMP: siehe 3.3.2). Auch hier erfolgt die Umsetzung bei pH 8 innerhalb von 20 Sekunden (Zeitfenster). Bei pH 6 und ausgeprägter bei pH 4 nimmt die Reaktionsgeschwindigkeit deutlich ab, bei pH 2 jedoch wieder zu. Die Zunahme der Reaktionsgeschwindigkeit der Reaktion von Melarsen Oxid mit den niedermolekularen Dithiolen bei niedrigen pH-Werten kann durch eine säurekatalysierte Veresterung - analog der Bildung von Carbonsäureestern - beschrieben werden. Mit zunehmenden pH-Wert würde sich dann die Geschwindigkeit der Reaktion verlangsamen. Die sehr viel größere Reaktionsgeschwindigkeit im Basischen läßt sich auf die deutlich höhere Nukleophilie des Thiolatanions (pKa 7-9) zurückführen.
Daß Oxytocin sich nicht analog den niedermolekularen Dithiolen verhält, muß auf einem strukturbedingten Unterschied beruhen. Denkbar wäre, daß Oxytocin im physiologisch nahen pH-Bereich eine Tertiärstruktur einnimmt, die die Ausbildung eines zyklischen Arsonigsäuredithioesters stabilisiert. Mit abnehmendem pH-Wert würde diese dann durch Konformationsänderungen des Peptidgerüsts destabilisiert werden.
|
|
|
|
|
2.3 Massenspektrometrische Charakterisierung der Reaktionsprodukte von Modellpeptiden mit Arsonigen Säuren |
|
|
2.1 Synthese von Derivaten der Arsonigen Säure |
Diese Seite wurde erstellt am 15. März 1998. Letzte Änderungen am 30. Juni 1998 durchPeter Happersberger.