|
|
|
|
|
1.5 Zielsetzung und Aufgabenstellungen der Arbeit |
|
|
1.3 Massenspektrometrische Methodenzur Charakterisierung von proteinchemischen Umsetzungen |
1.4 Bedeutung, Struktur und biologische Funktion des Makrophagen-Kolonie-Stimulationsfaktors (M-CSF)
Das Cytokin Makrophagen-Kolonie-Stimulationsfaktor (M-CSF) gehört zu einer Gruppe von 18 bekannten Glykoproteinen, die als hämatopoesische (blutbildende) Wachstumsfaktoren fungieren [94-96]. Als komplexes Proteinhormon stimuliert M-CSF die Proliferation und Zelldifferenzierung von Phagozyten [97] und ist für das Überleben und die Stimulation von Makrophagen und deren Vorläuferzellen essentiell [98,99]. So wird durch M-CSF die Fähigkeit reifer, einzelliger Phagozyten verstärkt, infektiöse Mikroorganismen und Tumorzellen zu eliminieren [100]. Derzeit werden therapeutische Anwendungsmöglichkeiten des M-CSF in klinischen Tests untersucht. Von potentiellen Nutzen wäre M-CSF z.B. für eine beschleunigte Wiederherstellung des normalen Blutbildes chemo- und strahlentherapeutisch behandelter Krebspatienten, sowie zur Bekämpfung von Immunschwäche-induzierten Pilzkrankheiten [101]. Ferner ergaben vorklinische Studien, daß die Behandlung mit M-CSF zu einer Absenkung eines erhöhten Cholesterinspiegels führt [102].
Des weiteren wird die biologische Funktion des M-CSFs bei pathologischen Prozessen untersucht. Infolge von Entzündungen erhöhen sich die M-CSF-Serumkonzentrationen, wie auch die der meisten anderen Cytokine. Die Tatsache, daß auch hohe M-CSF-Konzentrationen in Individuen ohne Entzündungen gemessen werden [101], deutet auf die besondere Rolle des M-CSFs in der normalen, nicht pathologischen Hämatopoese (Blutbildung) hin. Stark erhöhte M-CSF-Werte lassen sich auch in Geweben von Patienten nachweisen, die an Ateriosklerose [103], Autoimmunkrankheiten oder Mamma- [104] oder Ovarialkarzinomen [105] erkrankt sind. Die Ursachen dieser Erkrankungen sind zum gegenwärtigen Zeitpunkt erst unzulänglich geklärt, auch wird die Rolle des M-CSFs bei der Tumorgenese kontrovers diskutiert.
Weitere verwendete Bezeichnungen für M-CSF sind:colony-stimulating-factor-1 (CSF-1), macrophage growth factor(MGF), macrophage and granulocyte inducer (IM oder MGI-IM) [106].
1.4.1 Biologisch aktive Formen des M-CSF
Humanes M-CSF tritt als biologisch aktives Homodimer entweder als integraler Bestandteil von Zellmembranen oder in strukturell unterschiedlichen sezernierten Formen auf [107]. Dabei variiert das Molekulargewicht der nativen humanen M-CSF-Formen zwischen 68 und 160 kD. Humanes M-CSF wird in Zellen durch ein Gen codiert. Durch alternatives mRNA Splicing [108-110] entstehen drei unterschiedliche M-CSF Formen, deren cDNA sequenziert wurde. Die drei Formen werden als M-CSF (256 AS) [95], M-CSF (554 AS) [111] und M-CSF (438 AS) [108] bezeichnet (sieheAbb. 15). Bei allen drei Proteinen ist die Sequenz der ersten 149 N-terminalen Aminosäuren, sowie die Sequenz der 75 C-terminalen Aminosäuren identisch. M-CSF und M-CSF weisen jedoch ab AS 149 Insertionen bzw. Deletionen bei M-CSF [110] auf.

Abb. 15: Schematische Darstellung der drei alternativen Formen des sezernierten M-CSF. Die Sequenz des Monomers ist in Balkenform dargestellt; TM kennzeichnet die postulierte Transmembranregion. Das in dieser Arbeit verwendete rhM-CSF ist als Sequenzabschnit (4-221) des M-CSF gekennzeichnet.
In der vorliegenden Arbeit wurden alle Untersuchungen an einem rekombinanten humanen M-CSF (kurz rhM-CSF) durchgeführt. Dieses biologisch aktive, nicht glykosylierte Protein wurde als verkürzte Variante des M-CSF (AS 4-221) in Escherichia coli exprimiert und nach seiner Isolierung in vitro renaturiert [112] (siehe 1.4.3).
1.4.2 Bekannte Struktur- und Funktionsmerkmale des M-CSF
Für das M-CSF wurden eine Reihe posttranslationaler Modifizierungen, wie z.B. Glykosylierungen [113] oder die Addition von Chondroitinsulfat [114] nachgewiesen. Die gefundenen O- und N-Glykosylierungen befinden sich vorrangig im C-terminalen Sequenzbereich und sind für die biologische Aktivität in vitronicht notwendig [112]. Vermutlich dienen die posttranslationalen Modifikationen zum Schutz des M-CSFs vor raschem proteolytischen Abbau in vivo [106]. Für die Rezeptorbindung sind nur die N-terminalen Aminosäuren 4-149 relevant, denn sowohl die verschiedenen Splicing-Varianten, als auch das rekombinant hergestellte rhM-CSF sind biologisch aktiv. Die Aufklärung der Disulfidstrukturen am rhM-CSF mit massenspektrometrische Methoden [115] zeigte, daß sechs der neun Cysteine des rhM-CSF Monomers intramolekulare Disulfidbrücken (Cys7-Cys90, Cys48-Cys139 und Cys102-Cys146) ausbilden. Die restlichen Cysteine bilden intermolekulare Disulfidbrücken (Cys31-Cys31, Cys157-Cys157 und Cys159-Cys159) und verknüpfen somit die beiden Monomerhälften kovalent zum Homodimer. Unklar ist, ob homologe oder heterologe Cys159/Cys157 Disulfidbrücken vorliegen [115]. Eine Röntgenstruktur mit einer Auflösung von 2.5 Å für die Aminosäuresequenz 4-154 eines rekombinanten, in CHO (chinese hamster ovaries)-Zellen exprimierten, humanen M-CSF [116] bestätigt sieben der neun Disulfidbrücken, die innerhalb des gemeinsamen Sequenzbereichs liegen (siehe Abb. 16).

Abb. 16: Röntgenstruktur des verkürzten rhM-CSF; -Helices und -Faltblätter sind in der Ribbondarstellung zu sehen. Die Disulfidbrücken sind für beide Polypeptidketten des Homodimers eingezeichnet.
Aufgrund der hohen Flexibilität der C-terminalen Proteindomänen, ab etwa Aminosäure 150, ließen sich bislang keine Kristalle der längeren M-CSF Varianten züchten [106, 116]. Die dreidimensionale Struktur des unglykosylierten M-CSF (AS 4-154) (siehe Abb. 16) beinhaltet ein aus vier Helices bestehendes Bündel, das durch die intramolekularen Disulfidbrücken (s.o.) fixiert wird. DiesesVier-Helix-Strukturmotiv definiert eine ganze Familie von Proteinhormonen, der auch die meisten Cytokine angehören. Die Topologie der Helices (sieheAbb. 17) wird mit "up-up-down-down" bezeichnet und tritt als Strukturmotiv ebenfalls beim humanen Wachstumshormon(hGH), Granulocyten- (G-CSF) undGranulocyten-Makrophagen-Kolonie-Stimulationsfaktor (GM-CSF), sowie bei den Interleukinen 2, 4, 5 auf. Jedoch läßt sich keine Übereinstimmung zwischen den Aminosäuresequenzen des M-CSF und der genannten Proteinhormonen finden. In jüngsten MS-Untersuchungen zur Charakterisierung der Rezeptorbindungsbereiche von rhM-CSF wurde in unserem Arbeitskreis durch selektive chemische Modifizierungsreaktionen die A-Helix (Abb. 17) als interagierender Bereich mit dem Rezeptor identifiziert [117].
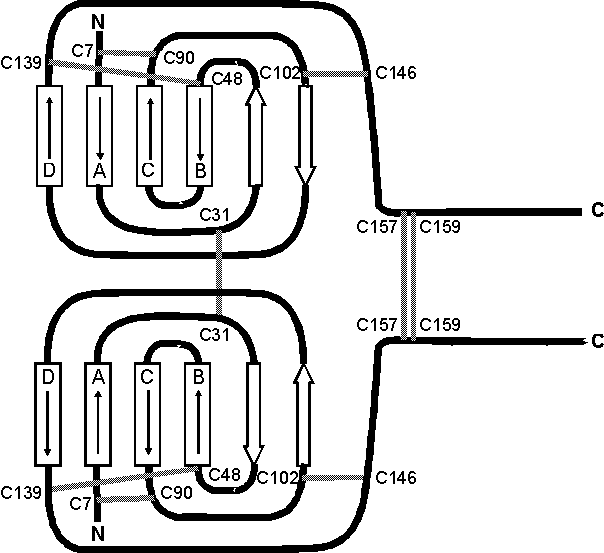
Abb. 17: Topologische Darstellung des rhM-CSF Homodimers zur Verdeutlichung des bei Cytokinen häufig anzutreffenden Strukturmotivs "up-up-down-down" der Helices A, B, C und D. Die Disulfidbrücken sind durch graue Balken gekennzeichnet. Die breiten Hohlpfeile symbolisieren die -Faltblätter.
1.4.3 Bekannte Rückfaltungreaktionen und Rückfaltungsintermediate von M-CSF
Bisherige Studien zur Rückfaltung von M-CSF erfolgten sowohl für die verkürzten, rekombinant hergestellten Formen der längsten M-CSF cDNA (siehe Abb. 15) als auch für rhM-CSF [116]. So isolierte Wilkins et al. monomere und dimere Faltungsintermediate des rhM-CSF (AS 4-153) [118]. Dieses Protein enthält nicht die zur Ausbildung der intermolekularen Disulfidbrücken notwendigen Cysteine Cys157 und Cys159. Die Bildung des biologisch aktiven Homodimers erfolgt durch die Disulfidbrücke Cys31-Cys31. Die Hauptfraktion der isolierten, durch Carboxamidomethylierung stabilisierten (siehe 1.1.3), monomeren Faltungsintermediate enthalten die intramolekularen Disulfidbrücken Cys7-Cys90, Cys48-Cys139 und Cys102-Cys146, während Cys31 carboxamidomethyliert vorliegt. Zu geringeren Anteilen konnten Wilkins et al. monomere Faltungsintermediate nachweisen, die zusätzlich an den Cysteine Cys102 und Cys146 carboxamidomethyliert waren. Das zusätzliche Auftreten dimerer Faltungsintermediate, mit den carboxamidomethylierten Cysteinen Cys102 und Cys146 jeweils eines Monomers, läßt darauf schließen, daß die Rückfaltung von M-CSF auf zwei unterschiedlichen Wegen (Mechanismen) erfolgen kann.
Untersuchungen von Glocker et al. [37] an dem in dieser Arbeit verwendeten rhM-CSF (AS 4-221) ergaben eine detailliertere Abfolge der Ausbildung der Disulfidbrücken während der Rückfaltung. Analog zu Wilkins et al. wurden die Faltungsintermediate durch Carboxamidomethylierung der freien Cysteine stabilisiert und über massenspektrometrische Peptide-Mapping-Analysen charakterisiert. Die Untersuchungen zeigten, daß zuerst die Ausbildung der intermolekularen Disulfidbrücken Cys157/Cys159 zum biologisch inaktiven Homodimer erfolgt. Die sukzessive Ausbildung der Disulfidbrücken Cys7-Cys90, Cys48-Cys139 und Cys31-Cys31 geht mit der zunehmenden biologischen Aktivität einher. Die Ausbildung der letzten Disulfidbrücke Cys102-Cys146 verleiht dem rhM-CSF Homodimer proteolytische Resistenz, was aus evolutionärer Sicht durchaus Sinn macht, da so eine Anreicherung proteaseresistenter Intermediate mit (eventueller) nicht nativer Struktur im Zellorganismus verhindert wird.
|
|
|
|
|
1.5 Zielsetzung und Aufgabenstellungen der Arbeit |
|
|
1.3 Massenspektrometrische Methodenzur Charakterisierung von proteinchemischen Umsetzungen |
Diese Seite wurde erstellt am 15. März 1998. Letzte Änderungen am 30. Juni 1998 durchPeter Happersberger.